What do we mean by regime shift? Distinguishing between extrinsic and intrinsic forcing in paleoecological data

Alistair W.R. Seddon
“Regime shifts” are abrupt changes in ecosystem function and state commonly observed in paleoecological records. Understanding whether they represent a linear response to an abrupt external forcing, or are the product of intrinsically mediated dynamics, remains an important distinction.
 |
|
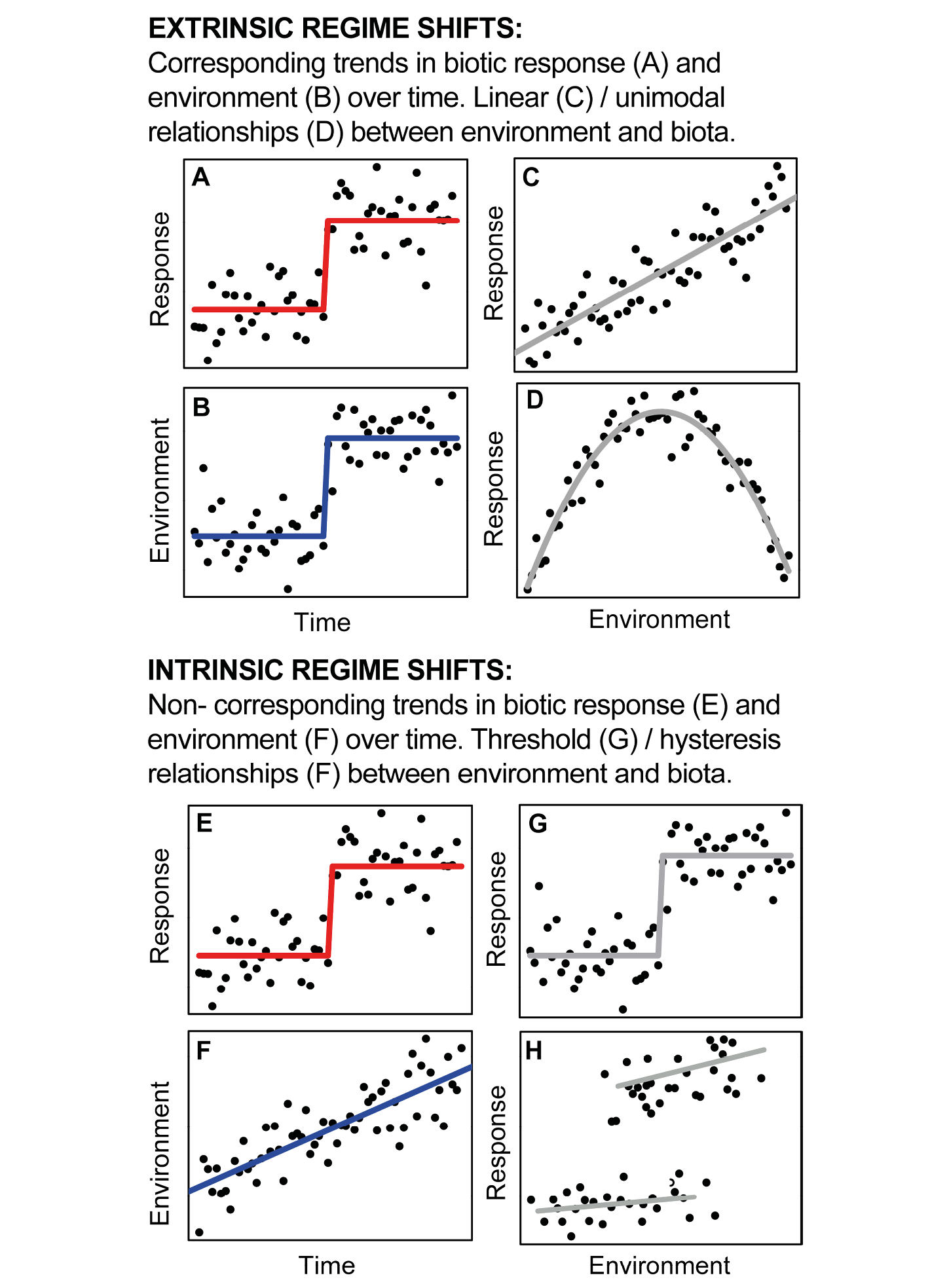
Figure 1: Framework used to identify intrinsic vs. extrinsic regime shifts (modified from Seddon et al. 2014). Equivalent trends in control and response variables, combined with linear or unimodal response functions represent extrinsic regime shifts (A-D). Gradual changes in control variables, combined with an abrupt shift in the response variable are more likely to represent an intrinsically mediated response (E-H). |
Ecological theory indicates that ecosystems can experience “regime shifts” – abrupt changes between two or more ecological states, each characterized by their own dynamics, stochastic fluctuations, or cycles (Scheffer 2009). The paleoecological record provides numerous examples of potential regime shifts, but interpreting the dynamics underlying these changes remains a major challenge. On the one hand, ecological regime shifts may represent linear responses to an external forcing; alternatively, they may be the result of a series of intrinsic ecological mechanisms (Fig. 1). Here, I provide a summary of some of my own recent work that aims to distinguish between extrinsically forced and intrinsically mediated transitions, noting that care must be taken not to conflate these different types of regime shifts when interpreting abrupt changes in the paleoecological record.
Theoretical framework
A theoretical framework for understanding abrupt ecosystem change was proposed by Williams et al. (2011), contrasting extrinsically forced against intrinsically mediated ecosystem responses to an environmental forcing (Fig. 1). Abrupt biotic responses as a result of changes in climate during the cooling into, and subsequent warming out of, the Younger Dryas were used as examples of extrinsically forced ecosystem change. In contrast, intrinsically mediated responses result from a combination of site-specific abiotic factors (e.g. soil characteristics, groundwater regime and physiography), or from local-scale biotic processes (competition, facilitation and disturbance) (Williams et al. 2011). Under these conditions, abrupt ecological changes can occur following only a gradual external forcing (e.g. Holocene transitions at the prairie-forest boundary in the North American Midwest).
One type of intrinsic regime shift that has become of particular interest to paleoecologists are fold-bifurcation-type transitions (Fig. 1h), in which positive feedbacks lead to sudden and abrupt transitions in response to slow forcing mechanisms. For example, in shallow lake ecosystems, theory indicates that transitions from clear to turbid waters are the result of a loss of system resilience through gradual nutrient loading (Scheffer et al. 1993). The feedbacks associated with maintaining a system in the turbid state mean that managing a transition back to the original state can be difficult to achieve. Because such “critical transitions” can occur with little apparent warning, understanding their mechanisms has become a major focus for the ecological science community. Furthermore, critical transitions and their associated loss of resilience have been proposed to have been observed a number of times in the paleoecological record.
One of the key insights presented in the framework presented by Williams et al. (2011) is that abrupt ecological responses can occur as a result from both linear and non-linear dynamics. This is important to recognize, because not all abrupt changes observed in the paleoecological record might represent intrinsic regime shifts. The challenge for paleoecologists is to develop the tools and techniques that can be used to disentangle extrinsic and intrinsic responses.
Distinguishing extrinsic and intrinsic regime shifts
One key challenge for studying regime shifts in paleoecological data is the unevenly spaced samples resulting from variability in sedimentation rates. A generalized non-linear least squares (GNLS) regression model can be used to account for this issue, which enables detection of changes in mean and variance between two variables, whilst simultaneously accounting for temporal dependence of data points which are unevenly spaced in time (e.g. Randsalu-Wendrup et al. 2012; Seddon et al. 2014). An additional advantage of GNLS is that a variety of trends can be implemented (e.g. linear, quadratic, logistic), meaning that trends in time series (i.e. ecological variable regressed against sample age) and response functions (ecological variable regressed against environmental driver) can be determined.
Seddon et al. (2014) used GNLS to distinguish intrinsic and extrinsic dynamics in a late-Holocene coastal-lagoon sequence from the Galapagos Islands. Following the framework from Williams et al. (2011), they used independent proxy data to reconstruct a range of habitat and coastal disturbance changes in the lagoon, and statistical modeling to describe the trends in both diatom assemblage and environment across major biotic transitions, and the response functions between these variables. Linear or unimodal relationships between control and response variables, in addition to corresponding trends in the two-temporal series, were assumed to represent extrinsic regime shifts, whilst a slow or gradual change in the control variable, corresponding to an abrupt response, would provide statistical evidence of the importance of intrinsic mechanisms (Fig. 1).
 |
|
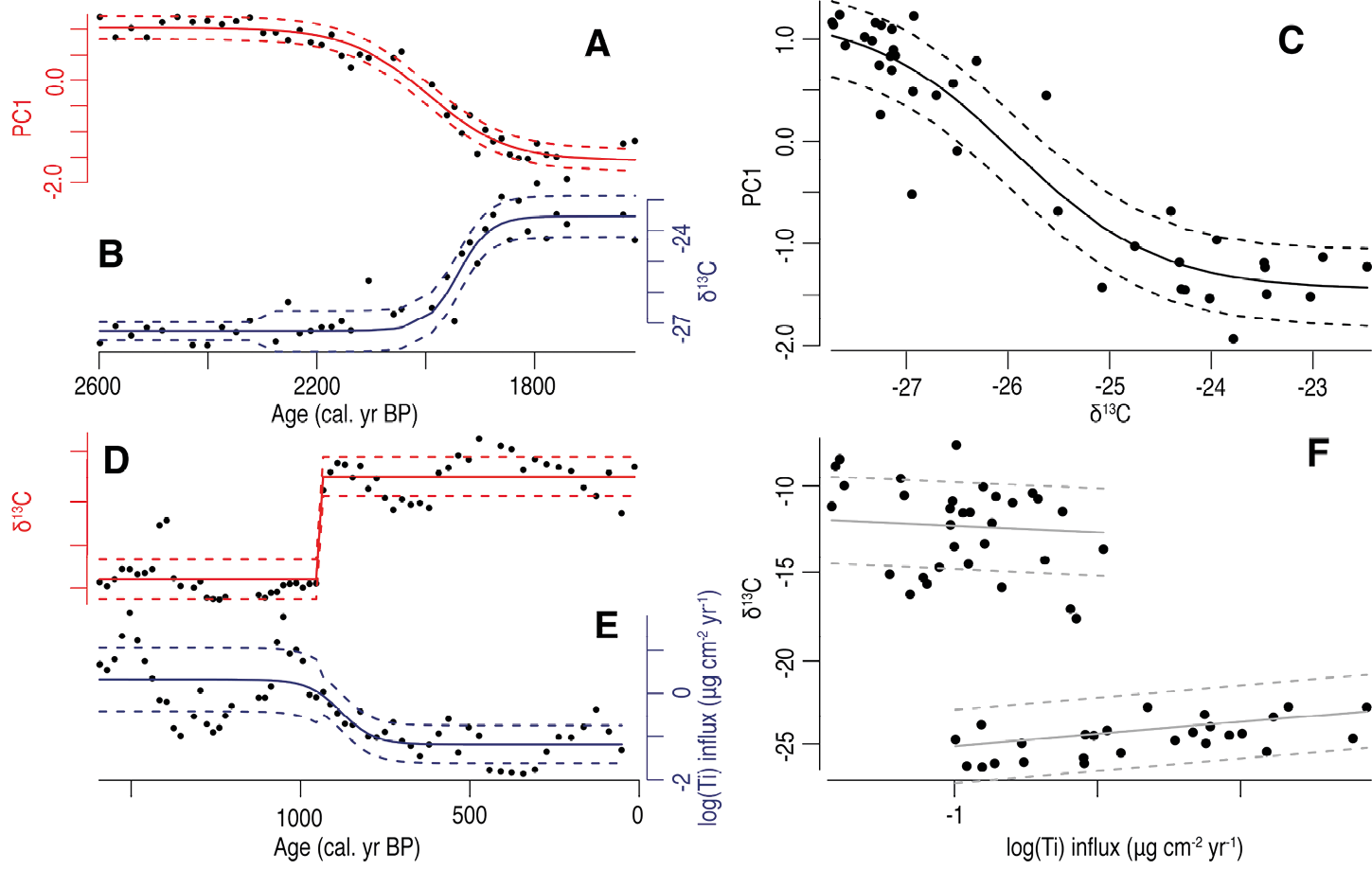
Figure 2: Evidence of extrinsic and intrinsic regime shifts at a coastal lagoon in the Galapagos Islands (modified from Seddon et al. 2014). (A, B) Example of an extrinsically forced regime shift. A change in habitat conditions (as detected by δ13C analysis of the bulk sediment) result in an equivalent response in the diatom assemblage data in the lagoon (as described by the first principle component of a PCA on the diatoms) with a curvilinear response function (C). (D, E) Potential evidence of an intrinsic regime shift. Minimal changes in the surrounding landscape (e.g. tidal disturbance events represented by geochemical data) result in an abrupt transition from mangrove (low sediment δ13C) to microbial mat (high sediment δ13C). Standard GNLS modeling was unable to adequately describe the relationship between the two variables, so the data were split at the major change in δ13C and two separate models were fitted to demonstrate the dual relationship with the control variable (F). |
The major diatom regime shifts observed in the lagoon showed the general characteristics of an extrinsic forcing (e.g. Fig. 2a-c). Given the fast generation times of the diatoms relative to the temporal resolution of the diatom record, perhaps this is not surprising (Hsieh and Ohman 2006), although critical transitions in diatom assemblages (i.e. intrinsic regime shifts) have been suggested elsewhere (Scheffer 2009; Wang et al. 2012). In contrast, the stable isotope data, representing transitions from a mangrove-dominated system to a microbial mat, appeared to occur at a time of limited salinity and other local environmental changes, hinting at evidence for intrinsic rather than extrinsic dynamics (Fig. 2d-f).
An alternative approach is investigating evidence for extrinsic responses from multiple paleoecological records. According to their framework, Williams et al. (2011) suggested that biotic responses to large extrinsic climatic drivers, such as those occurring during the last deglaciation, would be indicated by near synchronous responses across a large area. To investigate this, Seddon et al. (2015) applied a generalized additive mixed modeling approach to a series of ten high-resolution pollen records from Northern Europe. The modeling procedure tested whether a single smoother (i.e. pan-European, synchronous response), or multiple smoothers (i.e. individual site response) was the better explanation of the variance observed in the datasets. Results indicated that, during the early Holocene, vegetation changes provided evidence of similar behavior across sites, thus suggesting an extrinsic response.
Limitations and outlook
The studies outlined here represent just a few examples which have used quantitative approaches to identify and understand abrupt transitions. However, whilst these statistical methods are useful for identifying overall trends and associations within and between ecological and environmental temporal series, they do not fully characterize underlying mechanisms. Studies which can combine paleoecological data with process-based modeling are a challenge, but may be particularly useful for understanding the local-scale feedbacks, facilitation and competitive effects that mediate intrinsic responses in the paleoecological record (e.g. deMenocal et al. 2000; Jeffers et al. 2011).
A second limitation is that they are dependent on the availability of independent proxy data – using the framework outlined in Seddon et al. (2014) only works if all the key environmental forcing variables are available. Approaches such as early warning indicators, which use the underlying theory of critical slowing down to infer that fold bifurcations have occurred, have the potential to bypass this issue, but at present remain difficult to apply to paleoecological data on account of their sensitivity to unevenly spaced samples (Carstensen et al. 2013). Despite these limitations, statistical approaches can be used in conjunction with long-term ecological data in sediments to provide insights into the underlying dynamics of abrupt ecological transitions. Whether such transitions represent intrinsically mediated dynamics, or are the result of a linear response to an abrupt external forcing, remains an important distinction when interpreting abrupt changes in paleoecological records.
affiliations
Department of Biology, University of Bergen, Norway
contact
Alistair W.R. Seddon: alistair.seddon uib.no
uib.no
references
Carstensen J et al. (2013) Nature 498, E11-E12
deMenocal P et al. (2000) Quat Sci Rev 19: 347-361
Hsieh C-H, Ohman MD (2006) Ecology 87: 1932-1938
Jeffers ES et al. (2011) J Ecol 99: 1063-1070
Randsalu-Wendrup L et al. (2012) Ecosystems 15: 1336-1350
Scheffer M (2009) Critical Transitions in Nature and Society, Princeton University Press, 400 pp
Scheffer M et al. (1993) Trends Ecol Evolut 8: 275-279
Seddon AWR et al. (2011) PLOS ONE: e22376
Seddon AWR et al. (2014) Ecology 95: 3046-3055
Seddon AWR et al. (2015) The Holocene 25: 25-36