Multiple baselines for restoration ecology
Sandra Nogué![]() 1,2, L. de Nascimento
1,2, L. de Nascimento![]() 3, W.D. Gosling
3, W.D. Gosling![]() 4, N.J.D. Loughlin
4, N.J.D. Loughlin![]() 5, E. Montoya
5, E. Montoya![]() 6 and J.M. Wilmshurst
6 and J.M. Wilmshurst![]() 7,8
7,8
Recent work within restoration ecology has highlighted the importance of incorporating ecological history. Using three complementary examples from New Zealand offshore islands, the tropical Andes, and the Canary Islands, we discuss how restoration goals may be addressed using multiple baselines, or reference conditions, from long-term data.
The idea of using long-term ecological data to measure changes in biodiversity and to improve the effectiveness of conservation strategies has been widely discussed (Willis et al. 2010; Nogué et al. 2017). Importantly, the use of baselines, or reference conditions, has already been recognized in environmental policy such as in the conceptual framework for the 2019 Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES) global assessment report, and the United Nation decade on ecosystem restoration (2021–2030). It is, therefore, clear that there is now an urgent need to organize and standardize baselines if we want to implement successful restoration actions taking into account the complexity of managing both ecological and cultural systems (Jackson and Hobbs 2009; Nogué et al. 2021). We highlight three case studies to show how restoration goals may be addressed using paleoecologically informed baselines.
(1) Vegetation baselines for New Zealand offshore islands
New Zealand's offshore and outlying islands are important foci for biodiversity protection and conservation, with many supporting species that have been extirpated from the main islands (Bellingham et al. 2010). Most islands have been modified since initial human settlement began ca. 780 years ago, mainly by forest clearance and introduced mammalian pests (particularly rodents, cats, goats and pigs). Conservation efforts in the last 50 years have focused on eradicating alien species and replanting and restoring the "natural" vegetation. However, baselines have usually been informed by historic descriptions of vegetation and/or the composition of nearby forested locations.
Paleoecological records are recognized in New Zealand as an additional tool to document natural pre-human vegetation baselines and to help guide restoration plans (Wilmshurst et al. this issue). Long-term records have also been combined with local Māori (New Zealand's indigenous Polynesian population) knowledge, or mātauranga, to provide cultural vegetation baselines and to help inform biocultural approaches to island conservation (Lyver et al. 2015). For example, pollen and charcoal records showed how some forested ecosystems were cleared for Māori gardens, and local Māori communities now desire some forest to be restored to states that would provide a range of future benefits to their cultural, social, economic, and ecological needs (Lyver et al. 2015).
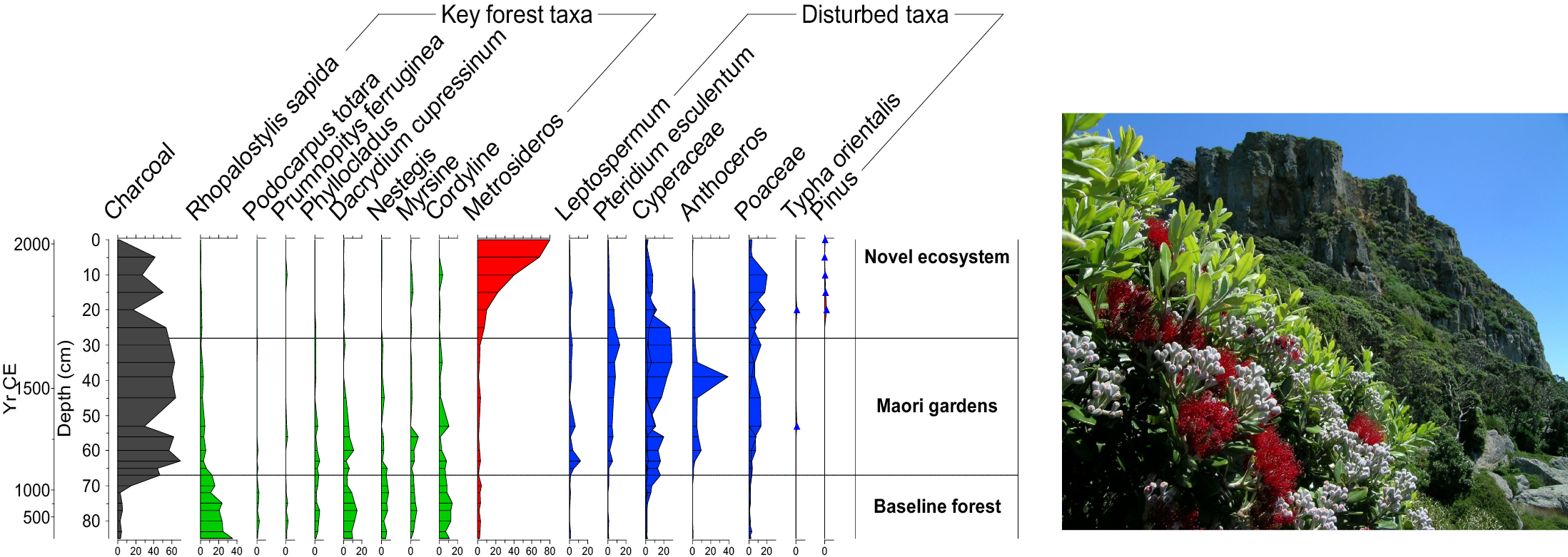 |
|
Figure 1: Example from Tawhiti Rahi Island of the Poor Knights Islands (Wilmshurst et al. 2014). Pollen data shows that the current vegetation composition is completely different from ancient ecosystems (photo credit: J.M. Wilmshurst). |
Sometimes paleoecological records can reveal surprises that overturn current ecological understanding. For example, the Poor Knights Islands in northeastern New Zealand are currently covered in a tall forest of native angiosperm trees dominated by Metrosideros. This vegetation composition has long been considered to be in a natural baseline state and is used to inform replanting of degraded islands in the region. However, a 2000 year record of pollen, charcoal and ancient plant DNA from a soil core on the island (Fig. 1) tells a different story (Wilmshurst et al. 2014). The forest was cleared by fire in the 13th century and gardened by Māori for ca. 550 years, followed by 180 years of forest succession after they ceased living there. The pre-human vegetation was completely different to today's forest, consisting of a diverse, conifer-dominated forest (e.g. Dacrydium cupressinum and Prumnopitys ferruginea) with an understory palm (Rhopalostylis sapida) and several other locally extinct tree taxa. Not only was Metrosideros unexpectedly uncommon in the past, but the pre-human baseline has no current analog on any northern New Zealand islands. This, and other paleoecological records from the region, demonstrate the power of the past to help inform future directions for conservation management.
(2) Examples from the tropical Andes
The Andean flank region has particularly caught the attention of researchers and policymakers due to its ecological value, and both its historical and current anthropogenic pressures (Cuesta et al. 2019). Paleoecological studies carried out in the tropical Andes have focused on using past environmental information to anticipate future scenarios of global change, including: (1) observing the synchronicity between environmental change and societal restructuring and adaptation in pre-Columbian populations (Gosling and Williams 2013); (2) pinpointing potential microrefugia locations based on the past dynamics of the Andean flag taxon Polylepis sp. (Valencia et al. 2016); and (3) highlighting shifted historical baselines such as those observed in the 19th century in locations with large pre-Columbian anthropogenic pressure that suffered depopulation following European arrival (Loughlin et al. 2018).
For example, fossil pollen showed a succession of Andean forest driven by changes in land-use. This forest succession was divided into five different vegetation baselines (Loughlin et al. 2018): pre-European (pre-CE 1588 indigenous occupation), successional (CE 1588–1718 European arrival/re-colonization), mature (CE 1718–1819 diminished population), deforestation (post CE 1819, re-colonization), and modern (industrial agriculture). These results were compared to a close site indicative of pre-human arrival vegetation (42 cal kyr BP; ancient), and showed the closest similarity with the post-abandoned mature state (Fig. 2). These new insights from sedimentary cores spanning the past three to four centuries show the potential to characterize meaningful restoration targets and reference ecosystems from paleoecological data (see Rull, this issue).
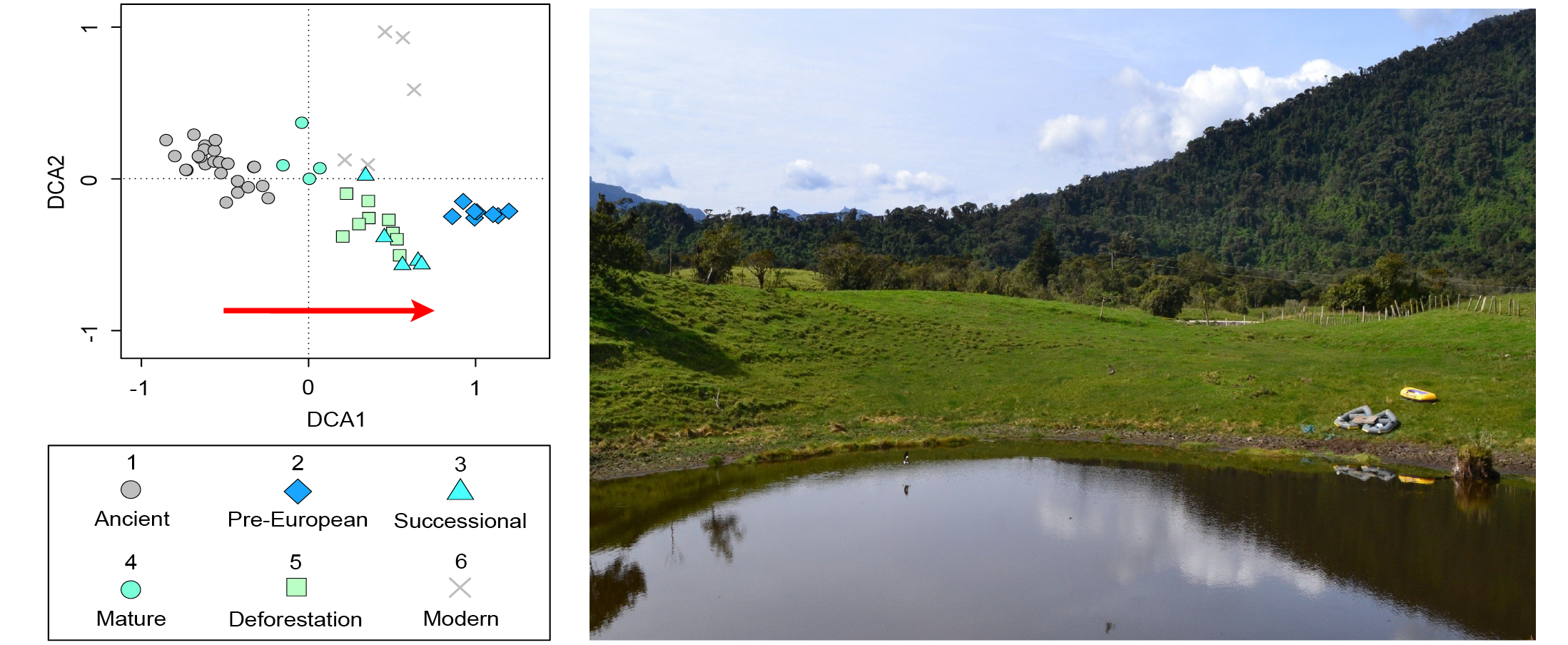 |
|
Figure 2: Detrended correspondence analysis (DCA) of pollen data from Andean montane forest grouped into time bins. Red arrow indicates an increasing signal of human impact. Replotted from Loughlin et al. (2018) (photo credit: N.J.D. Loughlin). |
(3) The thermophilous forest of the Canary Islands
The Canarian thermophilous woodland is among the most threatened ecosystems in the Canary Islands (del Arco et al. 2010). The remnants of these woodlands are currently occupying less than 10% of their original area and are undergoing several restoration actions. Describing baselines for these woodlands have proved challenging and is limited to the available ecological information from the few woodlands remaining on the islands.
On the island of Gran Canaria, there have been several restoration programs mostly focused on laurel forest remnants, since only 1% of its original distribution remains on the island today. One program included restoring the Laguna de Valleseco as part of the LIFE Laurisilva XXI project, with a key objective devoted to education (Cárdenes 1998). When this project was implemented, the pre-human baseline for the site was thought to be laurel forest with the presence of e.g. Ilex canariensis, Erica arborea, and Morella faya agreeing with the potential vegetation maps that predicted its presence in the area (del Arco et al. 2006). However, a paleoecological record carried out at the same site suggested a wider distribution of the thermophilous forest in the past, including this northern area. Fossil pollen records showed the unexpected presence of thermophilous species such as Juniperus and the date palm (Phoenix canariensis) until about 2300 years ago when there was an increase in fires, and grasses and shrubs spread over the area (de Nascimento et al. 2016). Before these new insights from the pollen record, Juniperus turbinata was considered a rare tree on the island (del Arco et al. 2006). This mismatch between data sources highlights the utility of combining paleoecological data into restoration planning, and improving understanding around past composition and occurrence of plant communities.
In Teno, on the island of Tenerife (European Union LIFE Habitat project), the reference condition was obtained by a combination of sources, including inventories of locally remaining Juniperus turbinata trees and patches of Juniper woodlands remaining in the archipelago (Tenerife and La Gomera), potential vegetation maps, toponymy (e.g. place name or geographic name), and historical and recent (elderly locals) accounts, to confirm past occurrence of the woodlands (Fernández-Palacios et al. 2008). Ten years after plantation, populations of J. turbinata are showing positive growth trends, with eight tree and shrub species surviving, growing, and some even showing signs of reproduction (flowers and fruits) (Rota et al. 2021).
Conclusions
With these three examples, covering ecosystems varying from high-mountains to islands, and from both hemispheres, we show the importance of incorporating paleoecological knowledge and other sources of long-term data in restoration programs. There are numerous examples of paleoecological research that have highlighted a priori misconceptions on the definition of the natural vegetation (Bush et al. 2014), and the ability of past environmental change data to characterize landscapes that are different to those of today (Rull et al. 2017). Paleo-informed baselines may provide a useful perspective on the variability of ancient systems, the magnitude of biodiversity change, novel ecosystems, and responses to successive waves of cultural transformation.
Acknowledgements
This work was supported by the Spanish Ministry of Science and Innovation (CGL 2009-10939), the Open University through a scholarship to N.J.D.L. (NE/L501888/1), and a NERC fellowship to E.M. (NE/J018562/1).
affiliations
1CREAF, Centre de Recerca Ecològica i Aplicacions Forestals, Bellaterra (Cerdanyola del Vallès), Catalonia, Spain
2School of Geography and Environmental Science, University of Southampton, UK
3Island Ecology and Biogeography Group, Instituto Universitario de Enfermedades Tropicales y Salud Pública de Canarias (IUETSPC), Universidad de La Laguna, Canary Islands, Spain
4Department of Ecosystem and Landscape Dynamics, Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, The Netherlands
5Geography & Environmental Sciences, Northumbria University, Newcastle Upon Tyne, UK
6Laboratory of Paleoecology, Geosciences Barcelona (CSIC), Barcelona, Spain
7Long-term Ecology Laboratory, Manaaki Whenua-Landcare Research, New Zealand
8School of Environment, University of Auckland, New Zealand
contact
Sandra Nogué: s.nogue creaf.uab.cat
creaf.uab.cat
references
Bellingham PJ (2010) N Z J Ecol 34: 115-136
Bush MB et al. (2014) Restor Ecol 22: 296-298
Cárdenes JL (1998) Medio Ambiente Canarias 11: 15-16
Cuesta F et al. (2019) Reg Environ 19: 1623-1642
del Arco MJ et al. (Ed) (2006) Mapa de Vegetación de Canarias. GRAFCAN, 552 pp
del Arco MJ et al. (2010) Biodivers Conserv 19: 3089-3140
de Nascimento L et al. (2016) Holocene 26: 113-125
Fernández-Palacios JM et al. (2008) Los bosques termófilos de Canarias (Proyecto LIFE04/NAT/ES/000064). Santa Cruz de Tenerife: Cabildo Insular de Tenerife
Gosling WD, Williams JJ (2013) Holocene 23: 1619-1624
Jackson ST, Hobbs RJ (2009) Science 325: 567-569
Loughlin NJD et al. (2018) Nat Ecol Evol 2: 1233-1236
Lyver POB et al. (2015) Hum Ecol 43: 681-695
Nogué S et al. (2021) Science 372: 488-491
Nogué S et al. (2017) Nat Ecol Evol 1: 0181
Rota F et al. (2021) Scientia Insularum 4: 27-44
Rull V et al. (2017) PAGES Mag 25: 82-83
Valencia BG et al. (2016) New Phytol 212: 510-522