Millennial to centennial changes in deep-ocean ventilation during the last deglaciation

Tianyu Chen1 and Laura F. Robinson2
The history of ocean ventilation helps to resolve timing and pathways of carbon transfer between the ocean and the atmosphere. Radiocarbon records reveal climate-linked, abrupt changes in the deep-ocean ventilation during the last deglaciation.
Radiocarbon as an ocean-ventilation proxy
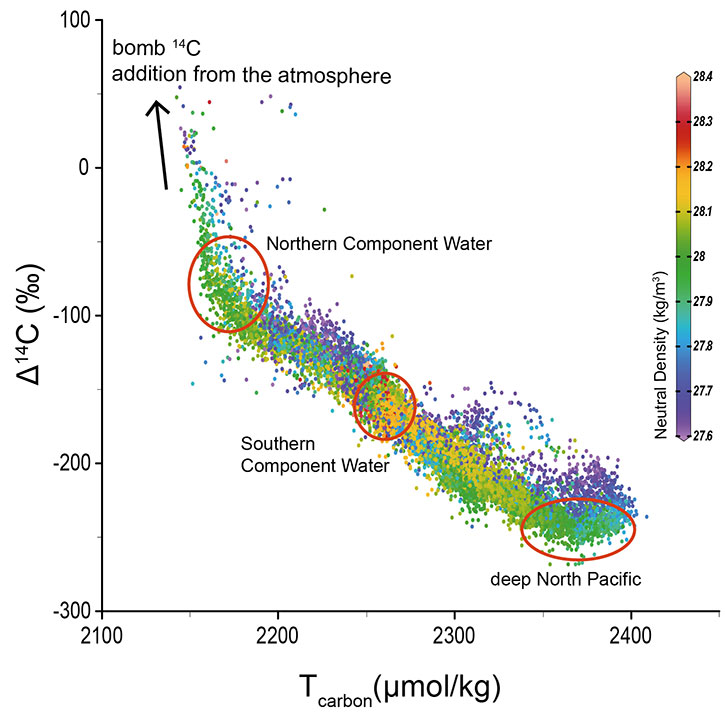
The transport of surface seawater to depths, known as "ventilation", represents a fundamental aspect of our climate system as it is tightly linked to the dynamics of overturning circulation and the global carbon cycle. Proxy-based reconstruction of past ventilation makes it possible to examine the interaction between the ocean and the atmosphere on long timescales when instrument records are not available. Among various proxies, 14C is one of the most sensitive for characterizing past deep-ocean ventilation. In the ocean interior, 14C is exclusively supplied through ventilation of the ocean from surface to deep, and it decays away with a half-life of 5730 years. Today, a large portion of 14C in the abyssal ocean (Fig. 1) is derived from the formation of North Atlantic Deep Water (NADW), with less than a third from overturning circulation in the Southern Ocean and biological remineralization (Broecker and Peng 1982). Since there is little deep convection in the present North Pacific, it contains the least ventilated waters at ~2 km depth (where the ventilation age is defined as the 14C age difference between the sample and the contemporaneous atmosphere) resulting from the slow transit of bottom water from the Southern Hemisphere. Along with the 14C-aging of the deep-water masses, there is a concomitant increase in the dissolved inorganic carbon inventory (~0.14 μmol/kg/year; Fig. 1) associated with the organic carbon respiration and calcite dissolution. As such, 14C is widely used as a semi-quantitative chronometer of ocean ventilation providing critical constraints on circulation dynamics and carbon storage in the past.
The reconstruction of past deep-water 14C signatures is not straightforward, as an independent age is required to correct for in-situ 14C decay in paleoarchives since their formation. Most deep-ocean radiocarbon records have been reconstructed using the fossilized remains of benthic foraminifera extracted from sediment cores (e.g. Skinner et al. 2010). Deep-sea scleractinian corals form a complementary emerging new archive that provides well-constrained, absolute-dated 14C records of the sub-surface ocean (e.g. Frank et al. 2004; Burke and Robinson 2012; Chen et al. 2015; Hines et al. 2015) since the Last Glacial Maximum (LGM).
Millennial-scale changes in ventilation
The low atmosphere CO2 concentration during the LGM is thought to be largely caused by coupled oceanic changes in ventilation and the biological pump. Indeed, compiled radiocarbon data suggest an ~700 14C-year increase in the average residence time of carbon in the deep ocean (> 1 km) during the LGM compared to the modern ocean, which allows the ocean to store more carbon (Skinner et al. 2017). So how did the deep ocean switch from an isolated, poorly ventilated LGM mode to the opposite in the Holocene? The key lies in the timing and pathways of the dissipation of 14C-depleted signatures of the deep ocean during the last deglaciation.
 |
|
Figure 1: Modern distribution of Δ14C and total carbon in deep waters of the global ocean (defined as those with neutral density >27.6 kg/m3). Note that nuclear bomb testing since the 1950s has dramatically increased the level of 14C in the atmosphere and subsequently deep waters in the North Atlantic. Figure was made with Ocean Data View, and the data are from GLODAP 2019. |
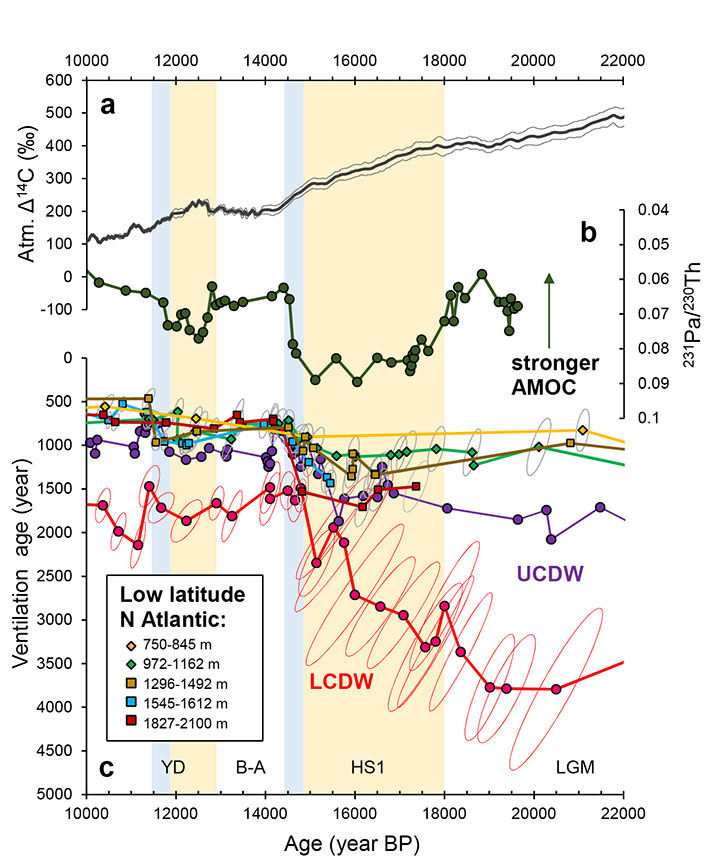
During the early deglaciation when the Northern Hemisphere was experiencing a cold stadial (Heinrich Stadial 1, HS1), the atmospheric CO2 concentration was increasing accompanied by a Δ14C decrease (Fig. 2; Reimer et al. 2013), implying mixing from a 14C-depleted CO2 source. Radiocarbon data from the upper and lower circumpolar deep waters (UCDW and LCDW) highlight converging trends in ventilation ages, indicative of increased Southern Ocean deep convection and air-sea exchange efficiency. This would have brought well-ventilated waters to depths and released 14C-depleted CO2 to the atmosphere (Anderson et al. 2009; Skinner et al. 2010; Burke and Robinson 2012; Chen et al. 2015).
Meanwhile, low-latitude North Atlantic deep waters at ~2 km remained poorly ventilated throughout HS1, some 700-800 years "older" than modern times (Fig. 2). This result is consistent with greatly reduced Atlantic Meridional Overturning Circulation (AMOC) during HS1 (McManus et al. 2004) that reduced the supply of 14C-enriched waters to the deep North Atlantic. In fact, the deep North Atlantic might have accumulated carbon during HS1 due to reduced ventilation (Menviel et al. 2018). Similar processes probably occurred during the Younger Dryas (YD) albeit with a smaller magnitude. Overall, larger 14C depth gradients are observed in the low-latitude North Atlantic during cold HS1 and YD compared to the Bølling-Allerød (B-A) warm event (Fig. 2), closely linked with changing production of the well-ventilated North Atlantic deep waters (McManus et al. 2004).
Centennial abrupt changes in ventilation
A particularly exciting aspect of the data being collected from deep-sea corals is the potential to reveal oceanic changes that occurred on centennial timescales. The transitions from HS1 to the B-A and from the YD to the Holocene are marked by abrupt increases in 14C of the low-latitude North Atlantic waters (Fig. 2), synchronous with warmings recorded in the Greenland ice cores and rapid increases in atmospheric CO2 concentrations (Chen et al. 2015). The coral 14C records from the Atlantic and the Southern Oceans also converge during these two transitions. This convergence likely reflects "flushing" events resulting from the abrupt resumption of the AMOC, which released a large amount of respired carbon from the deep ocean to the atmosphere (Chen et al. 2015).
 |
|
Figure 2: Ventilation age evolution of deep waters compared with other climate records. (A) Δ14C of the atmosphere (IntCal13, Reimer et al. 2013). (B) Sedimentary 231Pa/230Th ratios (an AMOC-strength index) from the subtropical North Atlantic (McManus et al. 2004). (C) Ventilation age reconstructed from deep-sea corals (UCDW and low latitude Atlantic; Burke and Robinson 2012; Chen et al. 2015) and benthic foraminifera (LCDW; Skinner et al. 2010). |
At the same time, boron-based pH reconstructions from Southern Ocean deep-sea corals further imply that these strong centennial ventilation events might be associated with delayed sea-ice advances at the beginning of the interstadial, allowing more efficient air-sea exchange (Rae et al. 2018). Multiple causes have to be invoked to account for the carbon release at the end of HS1 and the YD. Atmospheric δ13C records, for example, do not show the change expected from release of respired CO2 during the above two intervals, indicating increasing SST and/or increased land biosphere could also have played a role (Bauska et al. 2016). Moreover, the increased supply of nutrients to the surface is expected to stimulate primary productivity, enhancing the loss of alkalinity from surface waters and facilitating carbon release by increased overturning (Bronselaer et al. 2016). These puzzles are being addressed through increasing the resolution of data at these and other important oceanographic locations.
Concluding remarks
The upwelled 14C-depleted signatures from the abyssal ocean should eventually dissipate in the upper ocean and the atmosphere. Many uncertainties, unfortunately, still remain regarding intermediate water ventilation, with some 14C reconstructions closely tracking the evolution of atmospheric 14C while others showed extreme 14C depletions. Some studies further suggest that geological carbon addition to the water column could be an important component of the deglacial carbon cycle (e.g. Stott et al. 2019). In addition, consensus has not been reached regarding the magnitude and timing of 14C variability in the deep Pacific during the last deglaciation. Thus, the timing and pathways of deglacial oceanic carbon release have not yet been fully resolved. A growing understanding of 14C geochemistry in foraminifera and emerging deep-sea coral studies from the Pacific will provide a fresh look at these issues. Overall, 14C reconstructions of the deep ocean yield powerful constraints on ocean dynamics and carbon cycle during the last deglaciation.
affiliations
1School of Earth Sciences and Engineering, Nanjing University, China
2School of Earth Sciences, University of Bristol, UK
contact
Tianyu Chen: tianyuchen nju.edu.cn
nju.edu.cn
references
Anderson RF et al. (2009) Science 323: 1443-1448
Bauska TK et al. (2016) Proc Natl Acad Sci 113: 3465-3470
Broecker W, Peng T (1982) Tracers in the sea, Palisades, New York: 690 pp
Bronselaer B et al. (2016) Global Biogeochem Cy 30: 844-858
Burke A, Robinson LF (2012) Science 335: 557-561
Chen T et al. (2015) Science 349: 1537-1541
Frank N et al. (2004) Earth Planet Sc Lett 219: 297-309
Hines SK et al. (2015) Earth Planet Sc Lett 432: 46-58
McManus JF et al. (2004) Nature 428: 834-837
Menviel L et al. (2018) Nat Commun 9: 2503
Rae JWB et al. (2018) Nature 562: 569-573
Reimer PJ et al. (2013) Radiocarbon 55: 1869-1887
Skinner LC et al. (2010) Science 328: 1147-1151