Newly discovered Plio-Pleistocene sites in west Mt. Kenya: Potential tropical high elevation refugia?
The Central Highlands of Kenya (CHK) sites offer a unique opportunity to investigate evolutionary history during the Plio-Pleistocene period. Preliminary data indicate stable vegetation cover with continued hominin occupation of the sites, especially during extreme climates.
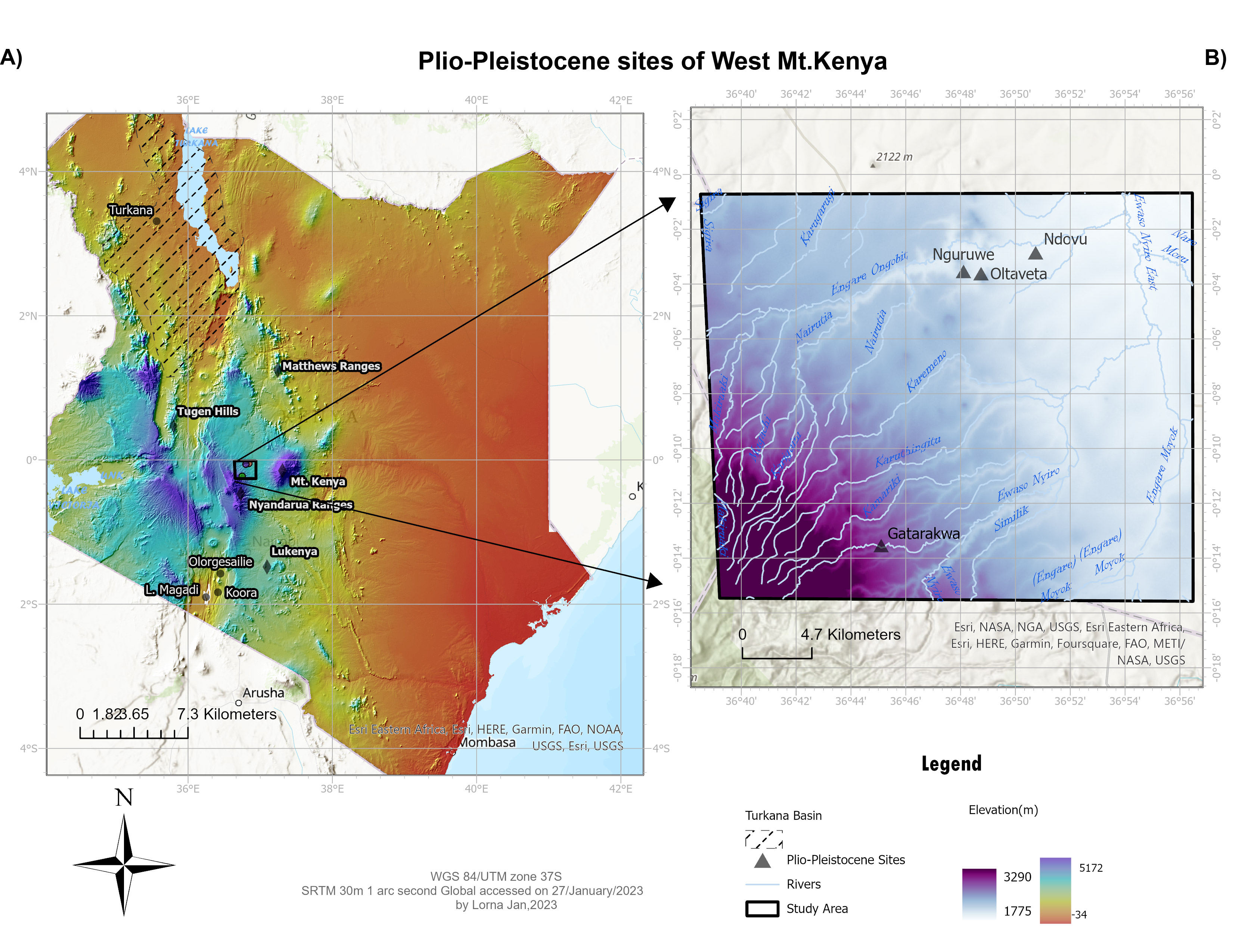
East Africa is renowned for its rich fossil and artefact record, especially on human evolutionary history, technological innovations, and advancement. Most East African prehistoric sites are found within, or in close association with, the East African Rift System (EARS). Here, we report on new fossil locality from the CHK that are unassociated with the EARS (Kirera et al. 2001; Malit et al. 2003). It is located at high elevation and surrounded by Mt. Kenya, the Aberdare Range, and the Mathews Range (Fig. 1). The mountain ranges influence local precipitation which differs from the country’s bimodal rainfall pattern (Nicholson 2017).
The CHK sites were discovered in 1999. Key fossil findings include remains of extinct megafauna; a fossilized tooth (lower p4) of a pig (Nyanzachoerus sp.); two kinds of extinct elephant (Deinotherium sp. and Anancus sp.); giant, long-horned wild cattle (Palerovis sp.); Carnivora; wild horses; small antelopes; and archaic Homo sapiens (Kirera et al. 2001; Malit et al. 2003). In addition, lithic technologies ranging from Early- to Late- Stone Age were also discovered. Based on the fossil evidence and the toolkits present, sites at the CHK span the last 5 Myr and may provide important information about a major extinction of African megafauna recorded at ca. 4.6 Myr BP (Faith et al. 2018).
To date, around 15 fossil-bearing sites have been identified at the CHK, providing a record that spans the earliest Pliocene through to the Holocene. Based on this fossil evidence and the transitional lithic technologies present, we conclude that the history of occupation at the sites is long, suggesting that CHK has been continuously resourceful and accessible to both fauna and humans for the Plio-Pleistocene (Kirera et al. 2001; Waweru, personal communication). Hence, CHK was occupied through periods of extreme climate variability recorded during the Pleistocene (Kaboth-Bahr et al. 2021). These attributes make CHK unique compared to other sites associated with the EARS, which more often represent intermittent occupation.
During the Plio-Pleistocene, as well as today, the CHK ecosystem was characterized by a habitat mosaic. The main goal of studying the past vegetation at the CHK is to understand the role played by the high-elevation sites of the CHK in the evolution of hominins and associated fauna and flora in East Africa during the Plio-Pleistocene. Specifically, this paper addresses the question of environmental dynamics attributed to variable wet–dry climates using palynological data.
Geological, paleontological and archaeological settings
CHK sites are located at high elevation associated with complex geological activities of the uplift and volcanism linked to the formation of Mt. Kenya to the east and the Aberdare Range to the west (Fig. 1). Radiometric dating (40Ar/39Ar) of volcanic tuffs from the CHK yielded ages of ca. 3 Myr BP, while biostratigraphic correlation indicates an age of ca. 5 Myr BP. Faunal fossils at the Nguruwe site include extinct pig Nyanzachoerus cf. kanamensis, which has been recorded in Pliocene sediments in the Turkana Basin and Tugen Hills (Bishop 2010; Hill et al. 1986). Remains of the extinct bovids Megalotragus spp. and Damaliscus spp. are also found at Ngobit and Oltaveta sites, and were previously recorded in Pleistocene deposits at the Lukenya sites (Faith et al. 2012; Potts and Deino 1995). Ngobit sites also preserve lithic technologies from Early-to Late Stone Age. One of the sediment layers at Ngobit was radiometrically dated, yielding ages between 614 kyr BP and 500 kyr BP, and containing hominin skull fragments, as well as Acheulean and transitional Acheulean to Middle Stone Age tools (Waweru, personal communication).
|
|
|
Figure 1: (A) Map of Kenya showing the Central Highland Kenya (CHK) locality, mountain ranges, and other comparable sites associated with the East Africa Rift System (Turkana Basin, Tugen Hills, Lukenya). (B) Map showing location of different CHK sites. |
Site significance
Highlands, among other zones such as coastal forests, lake margins and riparian habitats, are identified as potential tropical Quaternary refugia (Basell 2008; Joordens et al. 2019; Lahr and Foley 2016). Paleontological and archaeological evidence suggest that CHK sites were most likely refugia sites, being resourceful and habitable especially in events of extreme aridity in the East African region (Faith et al. 2012; Maslin et al. 2014). As the tropical African climate periodically shifted between humid and dry conditions, the CHK provided favorable and preferred habitats through orographic lift and topographic attenuation of orbitally forced climatic changes (Maslin et al. 2014). We hypothesize that the CHK biomes were resilient to remote climatic forces and, therefore, continuously habitable from the Pliocene to the Holocene period.
Vegetation data
This paper presents preliminary vegetation data inferred from phytolith analyses at several CHK sites. Phytoliths are silica plant cells’ casts formed in plants during growth. Upon plant decomposition, the cell casts are deposited in the soils and stay preserved for long periods of time. Their unique morphological features enable identification and classification of plant families (sedges, palms, and grasses) and taxa (woody, grasses, and herbaceous) (Piperno 2006). Phytoliths are reliable proxies to distinguish between C3 and C4 grasses, offering critical information on past temperature and precipitation (Fredlund and Tieszen 1997). C3 grasses are adapted to cool climates in tropical highlands, while in temperate regions they are even found at low elevations. C4 grasses constitute the highest proportion of low-elevation vegetation cover, with C4 tall grasses thriving best in warm and humid climates, while C4 short grasses are adapted to hot and dry climates (Piperno 2006). We processed and analyzed 20 samples collected from Pliocene and Pleistocene CHK sites to investigate if vegetation cover and structure differed between the two geological periods. We compared the vegetation data with the available faunal data to examine if both datasets reflect similar paleoenvironmental conditions at CHK
Significance of the vegetation data
Figure 2a shows the relative proportion of plants identified to family level (grasses, palms, and sedges) and taxa level (herbs, grasses, and woody taxa) using fossil phytolith, while Figure 2b are examples of the morphotypes identified. Phytolith data from the fossil samples reflect mixed wooded grasslands consisting of C3 and C4 grasses at CHK during the Pliocene and Pleistocene. However, the proportion of woody versus grass taxa varies across samples within the same age range. Some samples have a significant proportion of sedge and palm phytoliths, indicating that wetland habitats were dominant at CHK. Phytolith data from the modern soil samples (Fig. 2a) indicate mixed wooded grasslands, but with proportions varying across samples. This suggests vegetation cover has been stable and consistent on the CHK landscape since the Pliocene period. CHK sites, therefore, have been a suitable habitat for a variety of faunal species, including hominins.
 |
|
Figure 2: (A) Diagram showing relative morphotypes abundance and identified plant families using fossil phytoliths analyzed from CHK sites. Vertical axis shows the geological period brackets of the sites and the samples analyzed. The horizontal axis shows relative morphotypes abundance and the identified plants/taxa in different samples. (B) Microphotographs of some of the phytoliths identified (from top to bottom): bilobate from grass, globular ornate from woody taxa, and achene from sedges. |
While paleontological data at CHK sites indicates strong evidence of a megafaunal extinction at the Pliocene-Pleistocene boundary, stability of vegetation data shown here indicates that it is unlikely that changes in vegetation contributed to that extinction. Instead, the stable composition of the paleovegetation likely contributed to act as refugia hotspots which supported species survival long before the extinction. Ongoing research at the CHK is trying to refute this hypothesis. Paleoenvironmental information in the East African highland sites is currently lacking, or scarce. Therefore, more research is necessary to provide crucial information on faunal evolution history in this region.
affiliations
1Department of Archaeology, Max Planck Institute of Geoanthropology, Jena, Germany
2Department of Earth Sciences, National Museums of Kenya, Nairobi, Kenya
3Human Origins Program, Smithsonian National Museum of Natural History, Washington, USA
4Department of Anatomy, College of Graduate Studies, Midwestern University, Glendale, USA
5Department of Biomedical Sciences, Mercer University, Macon, USA
6Council on African Studies at the MacMillan Center for International and Area Studies, Yale University, USA
7Department of Anthropology, SUNY, Potsdam, USA
8Department of Geosciences and the Environment, Technical University of Kenya, Nairobi, Kenya
9Unaffiliated geologist, Nairobi, Kenya
10Department of Geosciences, Georgia State University, Atlanta, USA
11Unaffiliated naturalist, Nairobi, Kenya
contact
Rahab N. Kinyanjui: kinyanjui shh.mpg.de (kinyanjui[at]shh[dot]mpg[dot]de)
shh.mpg.de (kinyanjui[at]shh[dot]mpg[dot]de)
references
Basell LS (2008) Quat Sci Rev 27: 2484-2498
Bishop LC (2010) In: Lars W (Ed), Cenozoic Mammals of Africa. California Scholarship Online, 821-842
Faith JT et al. (2012) Palaeogeogr Palaeoclim Palaeoecol 361-362: 84-93
Faith JT et al. (2018) Science 362: 938-941
Fredlund GG, Tieszen L (1997) Palaeogeogr Palaeoclim Palaeoecol 136: 199-211
Joordens J et al. (2019) J Hum Evol 131: 176-202
Kaboth-Bahr S et al. (2021) Proc Natl Acad Sci 118(23): e2018277118
Kirera FM et al. (2001) J Hum Evol 40(3): A11-A12
Malit NR et al. (2003) Am J Phys Anthropol 120: 145
Maslin M et al. (2014) Quat Sci Rev 101: 1-17