Late Quaternary megafaunal extinctions in southern Patagonia and their relation to environmental changes and human impacts
Natalia A. Villavicencio
The chronology of megafaunal extinctions in southern Patagonia suggest that they are due to a combination of human impacts, and vegetation and climate changes. Most extinct megafauna coexisted with humans for a relatively long time, and died out at different times.
By the end of the Pleistocene, the world lost most of its large mammal species in what is known as the Late Quaternary extinction event (LQE; Martin and Klein 1984), the magnitude and timing of which differs among continents (Koch and Barnosky 2006). For more than five decades, the discussion about the possible causes of extinction has revolved around the human impacts caused by modern humans moving out of Africa, climate changes associated with the glacial-interglacial transition or combinations of both (Martin and Steadman 1999). South America was one of the most severely impacted continents, losing around 82% (53 genera) of all its large mammal species with average body mass exceeding 44 kg (Brook and Barnosky 2012). In South America, human arrival and late glacial climate changes occurred within a relatively short span of time, although marked regional differences are present in both the timing and direction of climate change. These regional differences provide opportunities to test possible synergistic effects of these changes in driving the LQE (Barnosky and Lindsey 2010).
The case of Southern Patagonia
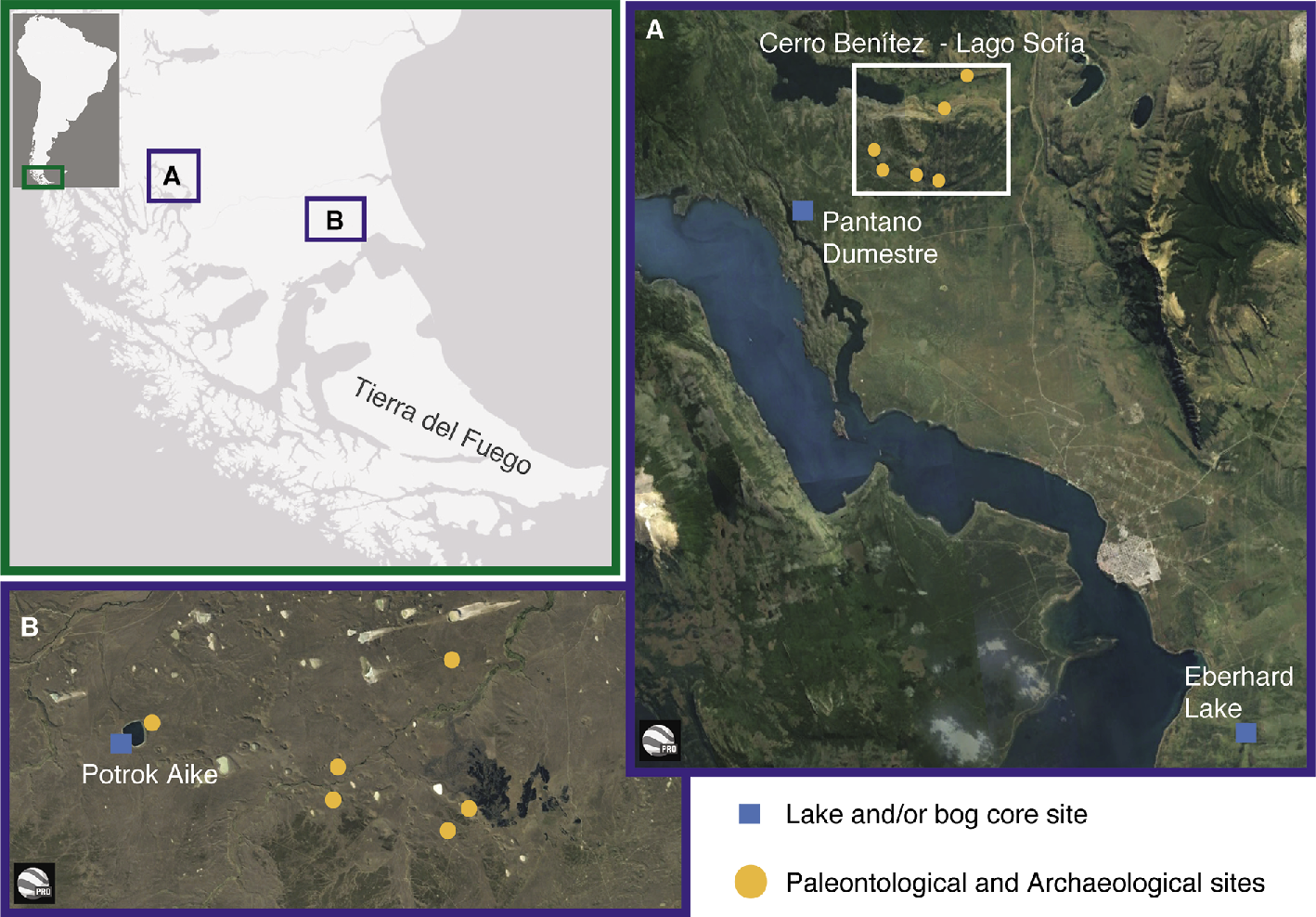 |
|
Figure 1: Map of southern Patagonia and location of study sites. (A) Última Esperanza, (B) Pali Aike. |
Several decades of research in caves and rock shelters in the area of Última Esperanza, Tierra del Fuego and Pali Aike (Fig. 1) have resulted in rich collections of extinct megafauna and evidence of the earliest human inhabitants in the area. Particularly, intense work in the Cerro Benítez and Lago Sofía area has produced dozens of radiocarbon dates on extinct fauna and helped establish one of the most complete chronologies of megafaunal extinction available from South America. Villavicencio et al. (2016) reviewed radiocarbon chronologies of megafaunal extinction and of human occupation from Última Esperanza, and the timing of major changes in climate and vegetation. Their main conclusions were (Fig. 2): First, mega-carnivores (Panthera and Smilodon) disappear slightly before the extinction of most of the herbivores, contrary to what is expected in a situation of trophic collapse. Second, Hippidion, Lama cf. owenii and mylodontids apparently coexisted with humans for hundreds of years (Hippidion and Lama cf. owenii) to millennia (mylodontids), ruling out a scenario of rapid overkill of these taxa by humans. Finally, Hippidion and Lama cf. owenii disappeared when cold grasslands were replaced by Nothofagus forest, followed by mylodontids disappearance, which occurred when the forests finally dominated the landscape.
Megafaunal extinctions in Última Esperanza
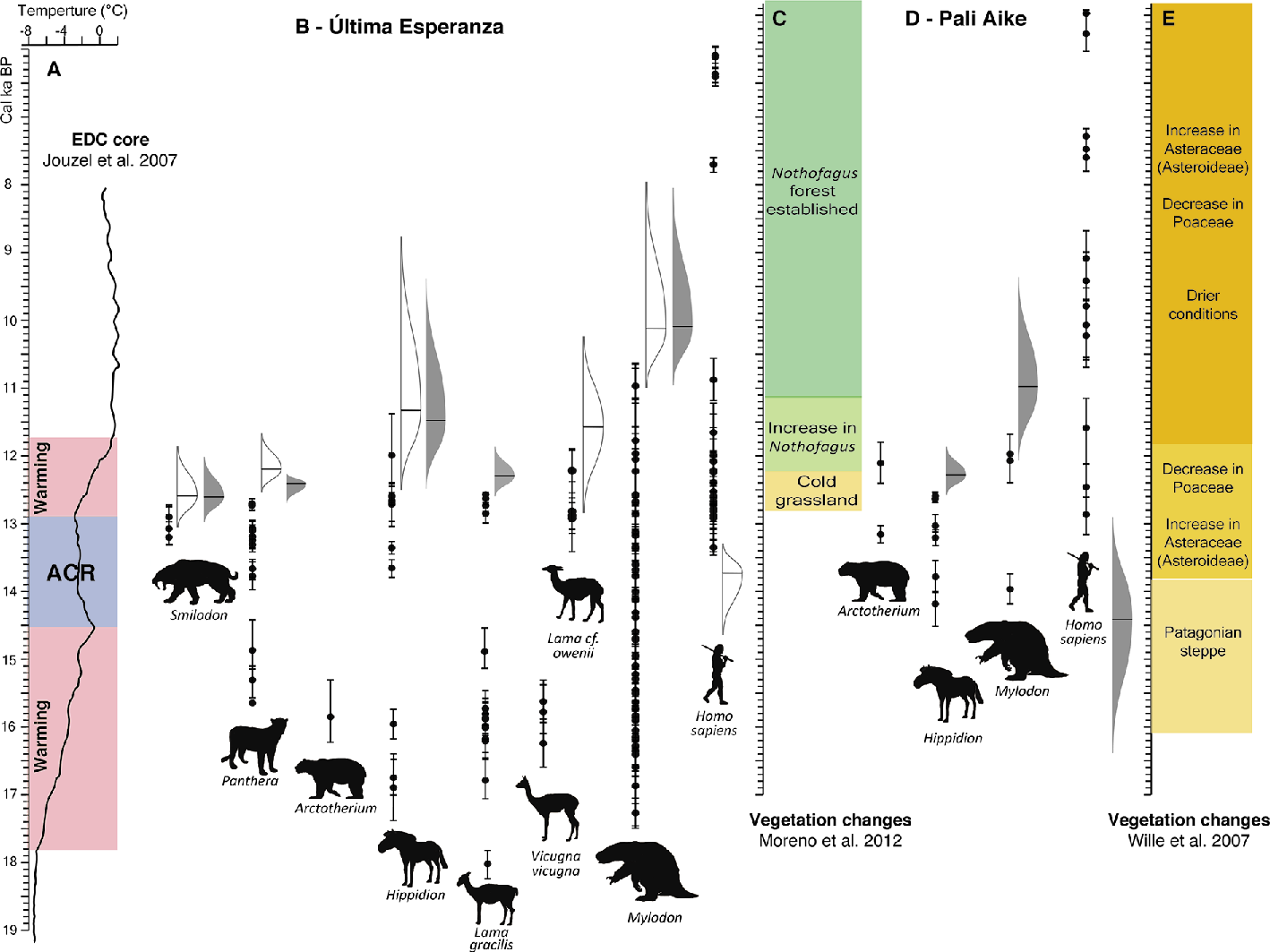 |
|
Figure 2: (A) Temperature record from the EPICA Dome C ice core, Antarctica. Blue bar: Antarctic Cold Reversal, red bars: warming events (Jouzel et al. 2007); (B and D) Chronology of megafaunal extinctions and human arrival in Última Esperanza (modified from Villavicencio et al. 2016) and Pali Aike; (C and E) Vegetation change estimations for Última Esperanza (Moreno et al. 2012) and Pali Aike (Wille et al. 2007). Black circles: direct calibrated (Calib 7.04; Stuvier and Reimer 2014) radiocarbon dates on extinct megafauna and human evidence. The GRIWM best-estimates of extinction timing (or timing of human arrival) are indicated by the unshaded (Villavicencio et al. 2016) and shaded (this work) normal distributions, with the 95% confidence band depicted by the areas around the mean (black line: most probable time of extinction). |
Over 45 new radiocarbon dates on extinct megafauna in Última Esperanza were recently published (Metcalf et al. 2016; Martin et al. 2015), making an update of previous analyses possible by combining these new dates with the Villavicencio et al. (2016) dataset of 62 high-quality radiocarbon dates. In the combined dataset, best estimates of local extinction times were obtained using the Gaussian-resampled, inverse-weighted method (GRIWM) of McInerny et al. (described in Bradshaw et al. 2012).
When the new dates are added, the general patterns of extinction as inferred from the GRIWM best-estimates appear almost the same as those calculated from the original 62 dates (Fig. 2). The extinction of Panthera and <Hippidion occurred earlier according to the new, combined dataset than according to the initial one. Lama gracilis, which was previously known by a single radiocarbon date, now has a more resolved record showing possible coexistence with humans for 900 to 1,200 years. This taxon disappears from the record between 12.6-12.2 ka BP, when the landscape was still dominated by cold grasslands. Four out of the eight taxa shown in Figure 2 become locally extinct during the warming phase following the Antarctic Cold Reversal cooling period recorded in the Antarctic EPICA Dome C ice core. Mylodon, and possibly Hippidion, disappear later from the record, between 12-10 ka BP.
Current archaeological evidence is scant and the number of dated human occupation events implies that Última Esperanza was visited on an ephemeral basis (Martin and Borrero, in press). However, the pattern of extinction remains evident, even after adding new radiocarbon dates to the analysis, and does not rule out the possibility of humans playing a role in driving some of these extinctions. Competition with humans could explain the disappearance of the large felids from the area between 700 and 2,000 years BP before some of the largest herbivores went extinct, as was proposed in Villavicencio et al. 2016. On the other hand, slow attrition of megaherbivores by human hunting, coupled with vegetation change, could also explain the later disappearance of mylodontids, horses and some of the extinct camels. Interestingly, human presence in the area fades at the same time of the last megafaunal extinction (Mylodon), reemerging more than 3000 years later.
Looking outside Última Esperanza: Pali Aike
Pali Aike is a volcanic field located east of Última Esperanza (Fig. 1). Archaeological research in the area dates back to 1936 (Bird 1988). Since then, numerous excavations have revealed the presence of extinct megafauna and humans during the late Pleistocene-Holocene transition (Martin 2013). This area is an interesting example to compare with Última Esperanza since it shares some of the same extinct taxa (Fig. 2) and is located in the same region (225 km to the east), but differs in the characteristics of human occupation and in the nature of both present-day vegetation and past environmental changes. Archaeological evidence for Pali Aike suggests more permanent human occupation in the area beginning ~12.9 ka BP (Martin and Borrero, in press). According to pollen records, Pali Aike was a cold Patagonian steppe dominated by grasses during the late Pleistocene, but transformed into shrubbier, less grass-dominated landscape between 13.9-11.9 ka BP, which has been interpreted as evidence for drier conditions (Wille et al. 2007). Southern beech (Nothofagus) forests found to the west never reached the area.
The megafaunal extinction record is less robust, when compared to results from Última Esperanza, as it consists of only 10 dates for three extinct megamammals. As shown in Figure 2, extinct fauna coexisted with humans for time periods ranging between 300 to 3,500 years. In general, the intervals of coexistence were shorter compared to Última Esperanza, which would be consistent with more permanent human presence in this area exerting greater pressures on megafauna. Coexistence between humans and Arctotherium is observed here, which has not been reported for Última Esperanza so far.
Resembling the extinction chronology in Última Esperanza, all three radiocarbon-dated taxa at Pali Aike disappeared from the record during the second warming period, after the Antarctic Cold Reversal, at a time when the landscape was becoming increasingly drier as inferred from pollen records (Wille et al. 2007).
Final remarks
The inclusion of 45 recently published radiocarbon dates in the chronology of extinction of Última Esperanza confirms the previously described general patterns and shows that a combination of human impacts and vegetation changes seem likely as the key drivers of megafaunal extinctions. Pali Aike shows interesting similarities with Última Esperanza - nonetheless more work is required, especially more radiocarbon dates on megafauna, in order to develop more robust conclusions.
acknowledgements
Thanks to Anthony D. Barnosky for insightful comments about this work.
affiliation
Department of Integrative Biology and Museum of Paleontology, University of California, Berkeley, USA
contact
Natalia A. Villavicencio: nvillavicencio berkeley.edu
berkeley.edu
references
Bird JB (1988) Travels and archaeology in South Chile. University of Iowa Press, 278 pp
Barnosky AD, Lindsey EL (2010) Quat Int 217: 10-29
Bradshaw CJA et al. (2012) Quat Sci Rev 33: 14-19
Jouzel J et al. (2007) Science 317: 793-796
Koch PL, Barnosky AD (2006) Ann Rev Ecol Evo Syst: 215-250
Martin PS, Steadman DW (1999) In: MacPhee RDE (Ed) Extinctions in near time. Springer, 17-50
Martin FM, Borrero LA (in press) Quat Int, doi: 10.1016/j.quaint.2015.06.023
Martin FM (2013) Tafonomía de la Transicion Pleistoceno-Holoceno en Fuego-Patagonia. Ediciones de la Universidad de Magallanes, 406 pp
Metcalf JL et al. (2016) Sci Adv 2, doi: 10.1126/sciadv.1501682
Moreno PI et al. (2012) Quat Sci Rev 41: 1-21
Stuiver M, Reimer PJ (2014) Calib 7.01 Radiocarbon Calibration