PAGES Magazine articles
Marie-Pierre Ledru1, A.C. Carnaval2, C.Y. Miyaki3 and “AF Biota” project participants4
The integration of paleoecological studies and genetic data from various Brazilian Atlantic forest organisms reveals three main regions of contemporary spatial distribution of genetic diversity related to late Quaternary shifts in monsoon activity.
 |
|
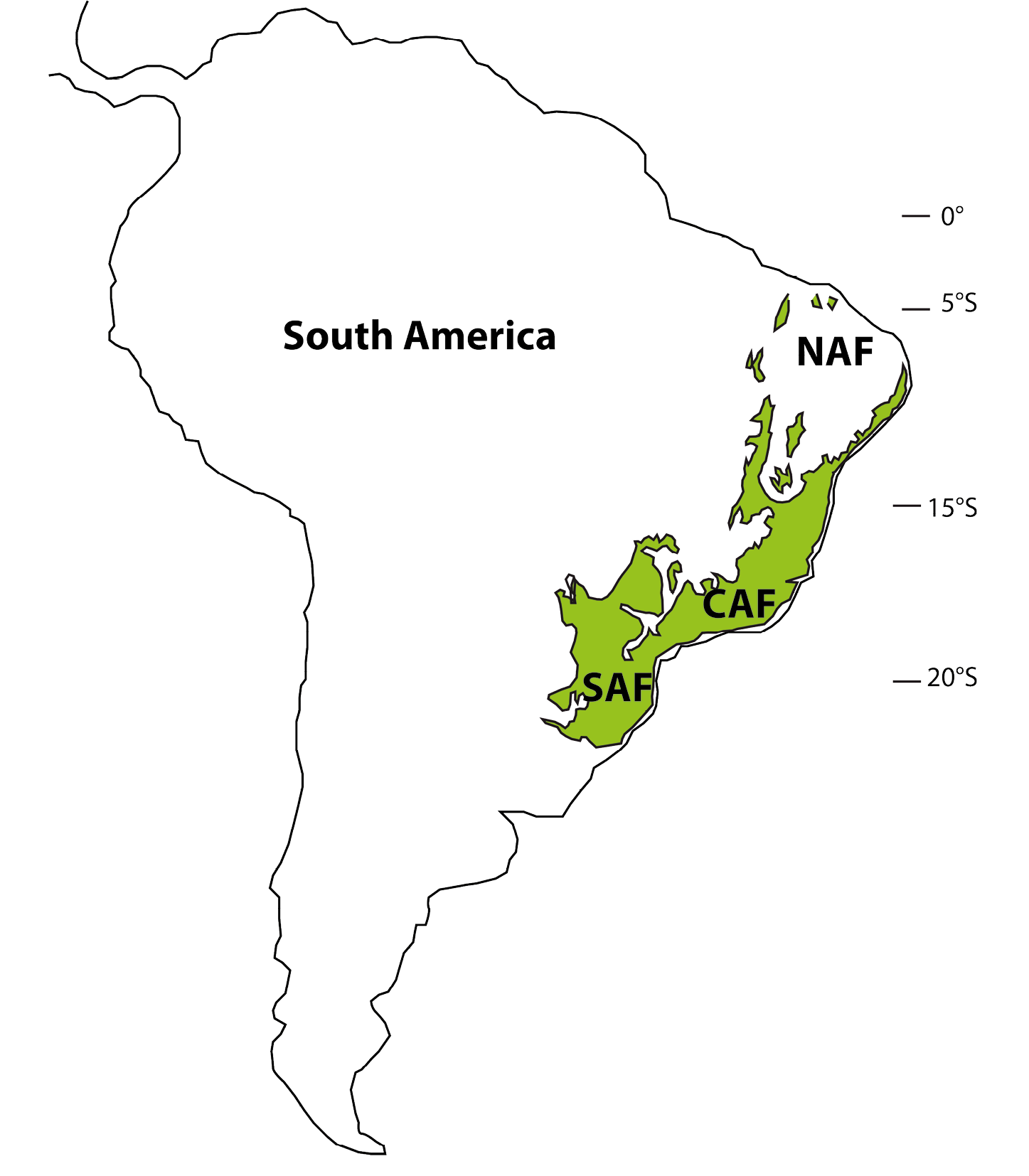
Figure 1: Map showing the modern distribution of the Brazilian Atlantic forest with three main regions. NAF: North Atlantic Forest; CAF: Central Atlantic Forest; SAF: South Atlantic Forest. |
The Atlantic forest is the second most biodiverse domain in Central and South America after the Amazonian rainforest. Its distribution today has been strongly reduced to less than 16% of its original cover because of intensive deforestation. The Atlantic forest covers a large region along Brazil’s 4000 km coast, from the Equator to 30°S (Fig. 1), and consequently is subject to a wide range of climatic conditions. These geographical characteristics, combined with a wide altitudinal range, resulted in one of the highest degrees of species richness and rates of endemism on the planet (Joly et al. 2014).
Three paleoclimatic regions
Modern regional climatic behaviors and past forest expansion and regression phases revealed by fossil pollen records (Ledru et al. 2016) define three main areas within the Atlantic forest: North Atlantic Forest (NAF) between 5° and 15°S, Central Atlantic Forest (CAF) between 15° and 23°S and South Atlantic Forest (SAF) from 23° to 30°S. NAF has a moist, cool, semi-deciduous forest restricted to coastal zones, lowland gallery forest and mountaintops. An evergreen forest was well developed during the late glacial instead of a deciduous forest (Fig. 2). CAF is composed of coastal forest patches of mainly dense evergreen forest, whereas more inland areas are occupied by semi-deciduous forests. The evolution of the CAF shows several phases of expansion and regression during the past 17 ka, which are linked to the precession cycle of the insolation (Ledru et al. 2009). SAF hosts Araucaria or mixed evergreen forest characterized by the presence of species adapted to cooler and wetter climates. The SAF expanded into the central region replacing the semi-deciduous forest during the late glacial, and since 3 ka is progressively expanding to the south (Fig. 2). This observation is in agreement with model predictions (Salazar et al. 2007). Two fossil pollen records from an interior northern site (Caço Lake, currently not forested) and from the central region (Colônia, currently forested) indicate that the expansion of the Atlantic forest cover was out-of-phase between these two regions - the northern site witnessed forest expansion from 17 ka to the beginning of the Holocene, while the central forest cover retracted (Ledru et al. 2016). Additional paleoecological datasets likewise indicate considerable expansion of the (nowadays small-ranged) northern forests during the deglaciation (17 to 12 ka; Wang et al. 2004).
Genetic diversity
 |
|
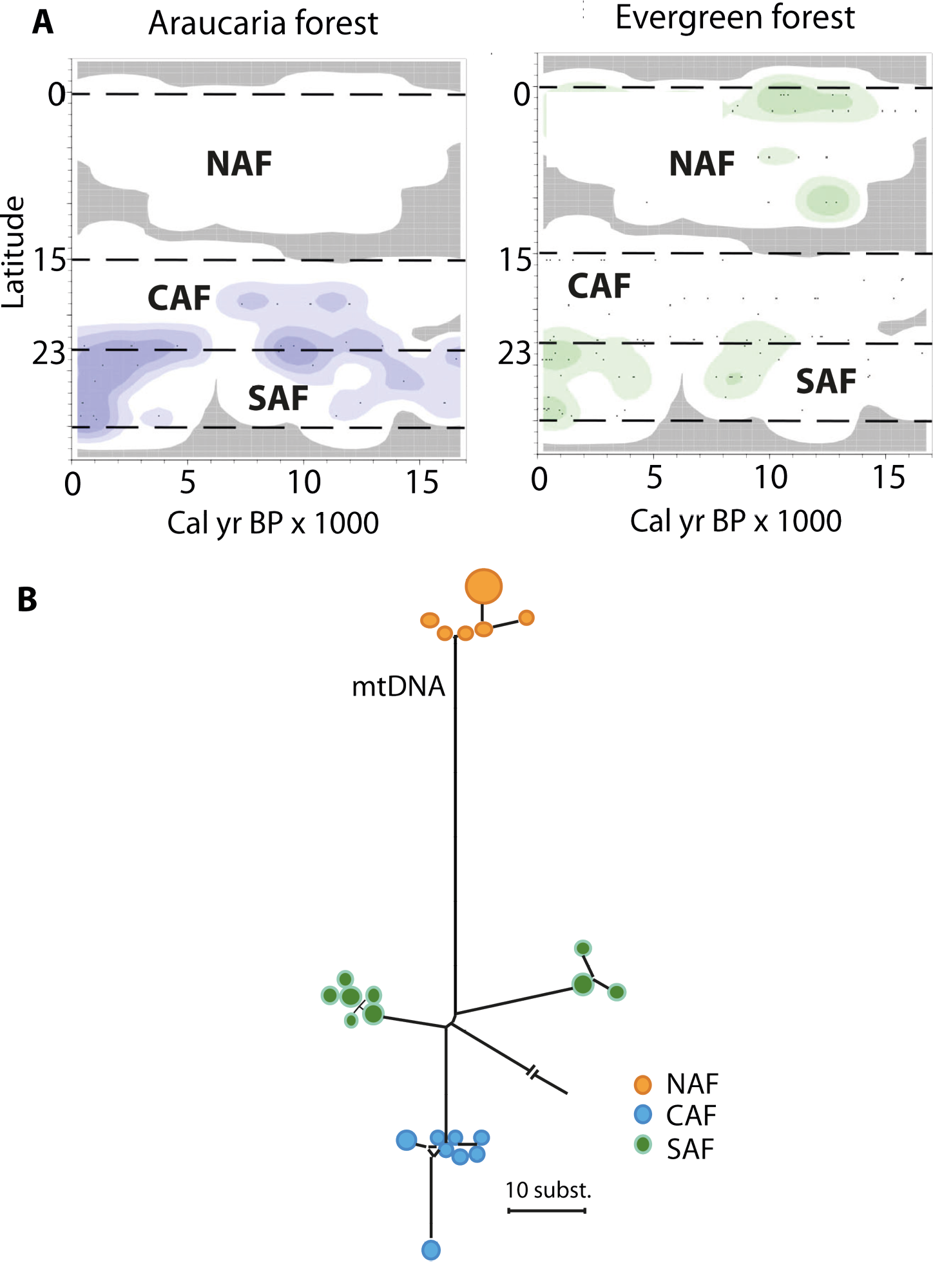
Figure 2: Paleoecology and biodiversity in the Brazilian Atlantic forest with: (A) Changes in the spatial distribution of groups of pollen taxa from Araucaria and evergreen forests as a function of time and latitude (modified from Ledru et al. 2016). Darker colors represent higher pollen concentrations. (B) Network showing relationships among mitochondrial DNA sequences of a bird species (modified from Cabanne et al. 2008). Each circle represents a sequence and their sizes are proportional to their frequency in the sample, the length of the lines represents the number of differences, and colors represent main areas (NAF, CAF and SAF). Note that two subgroups were distinguished within the SAF, but factors other than the monsoon shifts may have caused this separation. |
Comparative phylogeographic data from plants and animals provide a direct link between asynchronous Late Quaternary climatic shifts, the historical demography of forest-depended species, and genetic diversity patterns we observe today. If animal populations in the south have been tracking the Atlantic forest expansion in the late Holocene, then signatures of population expansion should be detected in DNA sequence data from multiple species. For instance, when a population becomes isolated, the succession of the generations within this population will generate new DNA imprints that will differ from the DNA imprints of the remote populations. On the other hand, populations restricted to northern, inland, highland sites must have been exposed to forest contraction over the same period and hence show genetic evidence of strong population bottlenecks that will reduce the diversity. NAF coastal populations, presumably exposed to permanently humid conditions throughout the last 21 ka (Carnaval and Moritz 2008; Carnaval et al. 2009), are expected to show high levels of genetic diversity due to climatic stability. Genetic data from coastal populations in the SAF confirm this hypothesis and show much lower levels of genetic diversity relative to NAF coastal populations, tied to genetic signatures of expansion and colonization that date post LGM. This pattern is detected in a vast array of forest taxa, including frogs, lizards, birds, bats, and plants (e.g. Cabanne et al. 2008; Carnaval et al. 2009; Fitzpatrick et al. 2009; Martins et al 2009; Ribeiro et al. 2010; Fig. 2). Post-3 ka colonization of the SAF is also seen in frogs and plants (Ramos et al. 2009; Carnaval et al. 2009). In turn, lineages restricted to mountaintop Holocene forest refugia in inland northeastern Brazil are narrowly endemic and show signatures of recent bottlenecks, presenting low levels of diversity (e.g. Ramos et al. 2009).
Climatic and diversity barriers
These asynchronous patterns of forest movements over the last 17 ka were tracked by various forest organisms, and largely determined contemporary spatial patterns of genetic diversity. Shift in monsoon activity, from north to south and south to north, driven by the precession cycles, led to significant differences in precipitation patterns (Cruz et al. 2005) and, thus, in the tempo and mode of rainforest contraction and expansion in lowland and mid-altitude tropical areas. This mechanism must have triggered strikingly different demographic trajectories between northern, central and southern species pools within forests distributed among this gradient. Such tracking of forested environments by whole tropical assemblages or species pools should largely explain contemporary patterns of genetic diversity in these tropical regions. In the Brazilian Atlantic forest, proposed forest refugia and areas of high genetic diversity and endemism match areas within the sea-saw monsoonal sector that were permanently moist over the last 17 ka. The three bioclimatic regions evidenced by phylogeography are located in the three paleoecological and paleoclimatic regions revealed by pollen records and related to past fluctuant monsoon activity at these latitudes.
The use of plant fossil information and genetic data from forest species to illustrate past climatic shifts and their associated responses from the fauna and flora of eastern South America question the timescale and pace of diversification, the role of refugia in tropical forest dynamics and the species-level tolerances to climate change.
affiliations
1Institut de Recherche pour le Développement (UMR ISEM), University of Montpellier, France
2Department of Biology, City University of New York, USA
3Departamento de Genética e Biologia Evolutiva, Universidade de São Paulo, Brazil
4Project “AF Biota” FAPESP (BIOTA 2013/50297-0), NSF (DEB 1343578) and NASA
contact
Marie-Pierre Ledru: Marie-Pierre.Ledru ird.fr
ird.fr
references
Cabanne SG et al. (2008) Mol Phylogenet Evol 49: 760-773
Carnaval AC, Moritz C (2008) J Biogeogr 35: 1187-1201
Carnaval C et al. (2009) Science 323: 785-789
Cruz et al. (2005) Nature 434: 64-66
Fitzpatrick SW et al. (2009) Mol Ecol 18: 2877-2896
Joly C et al. (2014) New Phytol 204: 459-473
Ledru M-P et al. (2009) Palaeogeogr Palaeoclimatol Palaeoecol 271: 140-152
Ledru M-P et al. (2006) Quat Sci Rev 25: 1110-1126
Ledru M-P et al. (2016) Biotropica 48: 159-169
Martins FM et al. (2009) BMC Evol Biol 9: 294
Ramos ACS et al. (2009) J Heredity 100: 206-216
Ribeiro RA et al. (2010) Heredity 106: 46-57
Sherilyn C. Fritz1 and Paul A. Baker2
Phylogenetic data have great potential for testing hypotheses about the nature and timing of geological and climatic events. We provide examples of how phylogenies have been used to constrain environmental history and propose the broader use of this approach.
Phylogeography investigates the evolutionary history (phylogeny) and spatial structure of groups of organisms (Avise 2000), often linking the history of biological lineages to Earth history. In this approach, similarities and differences in the genetic composition of various contemporary species or taxonomic groups are used to establish evolutionary relationships (phylogenetics), and this evolutionary history is time calibrated using dated fossils and molecular clock calculations. In phylogeography, the time-calibrated history of descent is subsequently related to coincident geologic events, such as mountain uplift, river formation and migration, glaciation, or climate change. These events may have created barriers to dispersal and fragmented populations (vicariance) or alternatively generated corridors for migration that increased gene flow among populations. The explosive development of genetic sequencing technologies has resulted in a proliferation of recent phylogeographic studies that reconstruct the evolution of life and link the history of diversification and speciation to the evolution of the environment.
This process of studying the role of geological and climatic processes in the evolution of ancient lineages also has been called “phylogeology” (Acosta et al. 2014). Such studies rely on a well-constrained history of geological and climatic events.
Often, the nature and timing of geological events is not well constrained, and contrasting scenarios of various aspects of Earth history have been proposed in the literature. We suggest that, in some of these cases, the most common direction of study, relating phylogenetic divergences to geological events, can be inverted, such that time-calibrated phylogenetic trees can be used to inform environmental history. We call this new integrated discipline of testing geologic scenarios with genetic data “geogenomics” (Baker et al. 2014). Here, we summarize a few recent studies that use phylogenetic data to constrain geologic history. In presenting these examples, we hope to encourage additional geogenomic investigations and increased communication among evolutionary biologists and geologists.
The evolution of riverine barriers
 |
|
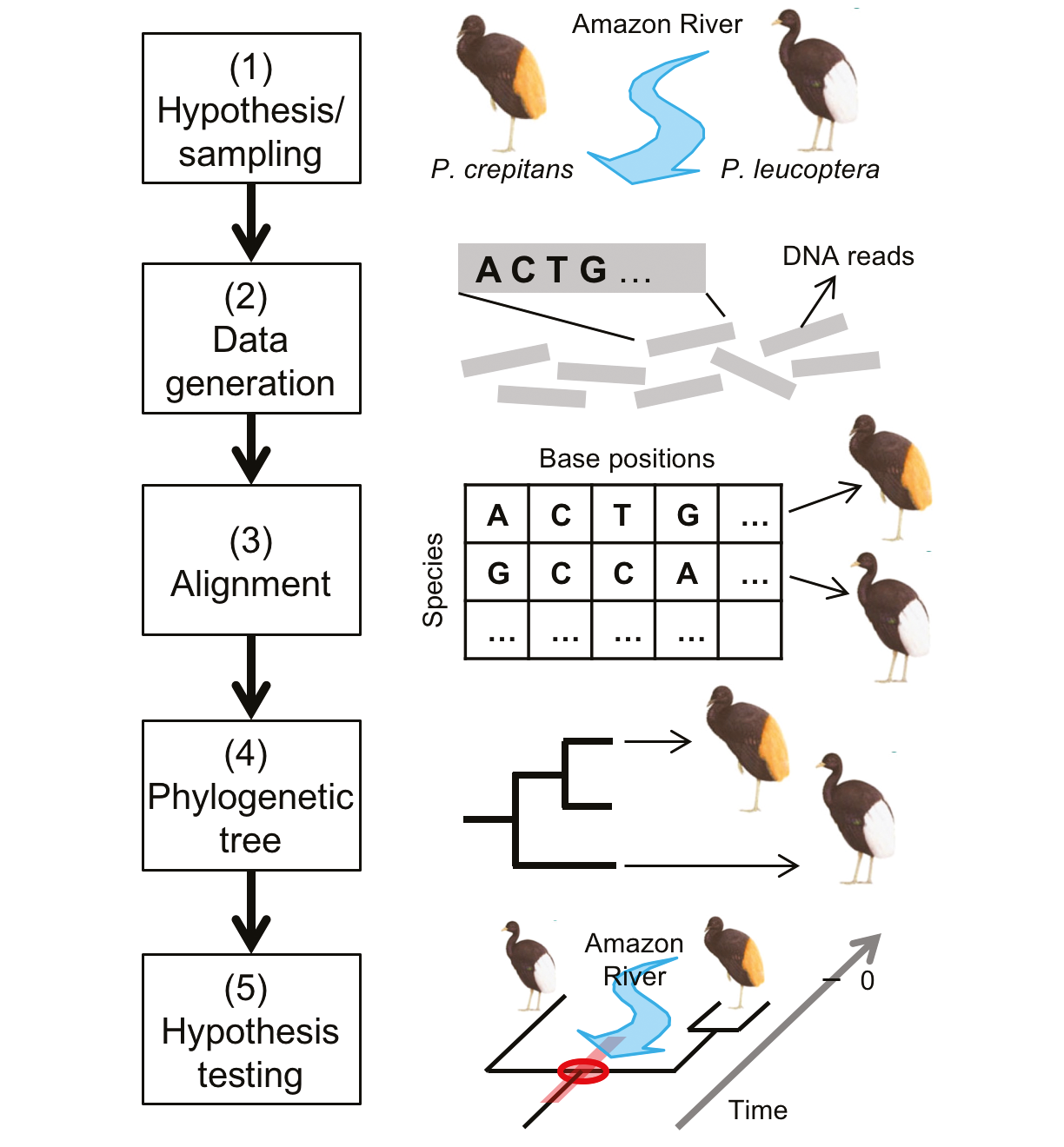
Figure 1: A schematic diagram showing how phylogenetic data on modern flightless bird populations has been used to test the timing of the formation of a river system. The phylogenetic tree defines the evolutionary relationship between bird species that occupy different river drainages in the Amazon, and the dated branching points in the phylogeny suggests events that split populations and led to genetic isolation and subsequent speciation. From Baker et al. 2014, reprinted with permission from Elsevier. |
A study by Ribas and colleagues (2013) of trumpeters, a group of flightless birds that exhibits a high degree of endemism, with different species occupying different drainage basins of the Amazon River system, first stimulated our interest in geogenomics and its potential for advancing understanding of Earth history (Baker et al. 2014). The timing of the development of the current trans-continental drainage pattern of the Amazon River is hotly debated, with estimates ranging from Miocene to Quaternary. Ribas and colleagues used a phylogenetic analysis of trumpeters to test different models of the geomorphic evolution of the river (Fig. 1). The analysis showed that diversification of trumpeters occurred relatively recently, between 3 and 0.5 Ma (millions of years ago), consistent with the more recent (Plio-Pleistocene) estimate for the establishment of trans-continental drainage. Moreover, the pattern of divergence between one clade (a group of organisms with a common ancestor) and another was used at finer resolution to constrain the ages of several major Amazon tributaries. Thus, the phylogenetic data were used to generate a testable hypothesis of geomorphic evolution.
The Great American Biotic Interchange
For many years, the Great American Biotic Interchange (GABI) between North and South America (Marshall 1988) was thought to be driven by the closure of the Isthmus of Panama and the generation of a continuous corridor of land between the two continents, dated at about 3 Ma. Yet recent geological studies (Montes et al. 2015) suggest a much earlier age for the closure (15-13 Ma), a hypothesis that is being contested in the geologic literature (O’Dea et al. 2016). Bacon and colleagues (2015) used molecular divergence dates, coupled with fossil data, to evaluate the timing of dispersal and vicariance in several different groups of organisms. Their analysis suggested a dramatic increase in the number of terrestrial taxa migrating between the continents about 3 Ma, but it also indicated increased migration during two earlier periods of the last 23 Ma, consistent with an earlier date for a land-based connection between North and South America. Similarly, marine organisms showed divergence in genetic structure consistent with an earlier separation of eastern and western populations by an emergent land bridge.
The hypothesis of an earlier (Miocene) closure of the Isthmus of Panama also has raised the question of whether the 3 Ma date for GABI derived from paleontological data is robust and also whether a different environmental event facilitated an increase in bi-directional exchange. One possibility is that faunal exchange was enabled by the expansion of savanna in the Central American corridor, caused by altered atmospheric circulation patterns associated with the onset of Northern Hemisphere glaciation. This hypothesis was proposed decades ago by several vertebrate paleontologists who first studied GABI, because many of the mammals involved in the exchange are adapted to savanna (Webb 1978). Bacon and colleagues (2016) conducted a subsequent phylogenetic analysis of mammals involved in GABI and calculated that the majority of changes in genetic structure between the continents occurred between 4 and 3 Ma. While this analysis does not address the nature of the environmental trigger, it does confirm paleontological estimates of the timing of major biotic exchange.
Glacial refugia
 |
|
Figure 2: (A) The genetic diversity (chloroplast DNA haplotypes) of white spruce populations across northern North America. High genetic diversity suggests population persistence for long periods, spanning back through glacial times. (B) Isolated spruce populations, similar in density to the refugial populations envisaged during intervals of North American glaciation. From Hu et al. 2009, reprinted with permission from the Ecological Society of America. |
Molecular genetic studies have revolutionized our understanding of the dynamics of plant populations following the retreat of continental ice sheets and montane glaciers at the end of the Pleistocene. In the Northern Hemisphere, paleoecological studies have used transects of pollen data to establish rates of migration and range expansion of tree populations that were thought to have survived glacial intervals in regions to the south and then migrated northward as the ice sheet retreated and climate warmed (Davis and Shaw 2001). More recently, however, population genetic studies of northern tree populations (Hu et al. 2009) have suggested that some species, such as white spruce, survived in refugia as small populations in areas adjacent to the ice sheet and subsequently expanded from these local areas to establish their current biogeographic distribution (Fig. 2). These new data have significant implications for calculations of potential tree dispersal rates and evaluating biological resilience to climate change. Early pollen analyses from Patagonia (Markgraf 1993) proposed that southern beech, Nothofagus, survived during glacial times in refugia on the now submerged coastal lowlands, rather than migrating long distances from the mid-latitudes, but this conclusion was controversial when originally proposed. Yet recent population genetic data (Premoli et al. 2010) indicate that species of Nothofagus persisted during glacial intervals both in ice-free regions of Patagonia, as well as in periglacial areas. Thus, a growing body of phylogenetic data indicates that in many regions small refugial tree populations persisted on the landscape through intervals of unfavorable climate.
Prospects for the future
The examples provided above suggest the potential for phylogenetic reconstructions to be used in generating or testing hypotheses about geological and environmental history. Doing so necessitates a well-developed understanding of the assumptions and uncertainties involved in phylogenetic reconstruction, just as biogeographic calibration of phylogenies based on geological data requires explicit accommodation of dating errors and an understanding of the mode and rate of the geological processes involved (De Baets et al. 2016). This need for a bi-directional flow of information suggests the value of expanded collaboration between geologists and evolutionary biologists in exploring the linkages between the evolution of life and the physical environment, and in furthering our understanding of the vital linkages between the biosphere and geosphere.
affiliations
1Department of Earth and Atmospheric Sciences, University of Nebraska - Lincoln, USA
2Earth & Ocean Sciences Division, Duke University, Durham, USA
contact
Sherilyn Fritz: sfritz2unl.edu
references
Acosta MC et al. (2014) Geobiology 12: 497-510
Bacon CD et al. (2015) PNAS 112: 6110-6115
Bacon CD et al. (2016) Geol Soc Am Bull 44: 375-378
Baker PA et al. (2014) Earth Sci Rev 135: 38-47
Davis MB, Shaw RS (2001) Science 292: 673-679
De Baets K et al. (2016) Phil Trans R Soc B 371: 20160098
Hu FS et al. (2009) Front Ecol Evol 7: 371-379
Markgraf V (1993) Palaeogeog Palaeoclimatol Palaeoecol 102: 53-68
Marshall LG (1988) Am Sci 76: 380-388
Montes C et al. (2015) Science 348: 226-229
O’Dea A et al. (2016) Sci Adv 2: e1600883
Premoli A et al. (2010) Palaeogeog Palaeoclimatol Palaeoecol 298: 247-256
Anneli Ekblom1 and Lindsey Gillson2
Cultural landscapes are enormously biodiverse. Presently, customary management of cultural landscapes is diminishing, resulting in loss of biodiversity and landscape heterogeneity. Here, we stress the role of paleoecology and other forms of paleoscience in restoring productive and biodiverse landscapes.
Extinction rates today rival the five major previous extinction events in Earth’s history. The biggest loss of biodiversity is due to land-use change, for example the conversion of forests, grasslands and other habitats through cultivation and urbanization, as well as the degradation and fragmentation of habitats (MA 2005). Stressed and fragmented ecosystems and populations are also less resilient to changing climate and other impacts such as pollution and invasive species. Restoration of ecosystem function, resilience and adaptive capacity is equally as important as maintaining species richness; therefore, different land-management options need to be considered. Intensive agricultural systems provide little wildlife habitat, whereas many land areas under traditional management (e.g. small-scale, low-intensive agricultural practices, referred to hereafter as customary) are significant havens for biodiversity (Fischer et al. 2012; Gillson 2015). Many cultural landscapes are biodiverse and are considered as important conservation targets (Agnoletti 2014). For example, cultural landscapes are now part of the International Union for Conservation of Nature protected areas classification. However, many customary management techniques have been eroded due to socio-economic pressures, with a subsequent loss of biodiversity and landscape heterogeneity. There is therefore a need to review customary management practices and the landscapes they produce, and to revive and reinforce the adaptive management techniques that are embodied in traditional land-use systems.
A role for paleoecology in landscape management
There is a role for paleoecology and other forms of long-term science in restoring productive, biodiverse landscapes. In many areas of Europe, rural land abandonment has led to the degradation of semi-natural woodlands, meadows and heathlands due to the discontinuation of customary practices (Weissteiner et al. 2011). Many species of high conservation value are specifically associated with these cultural landscapes and require a continued and active management. In Europe, 55 of the 231 listed habitat types of European interest depend on, or benefit from, continued customary practices (EEA 2011).
 |
|
Figure 1: Oakmeadow, central Sweden, is regularly grazed by cows to maintain the open meadow vegetation. Photo: Siri Pettersson. |
Paleoecological and historical studies have consistently shown a decrease in biodiversity associated with the loss of customary land-management practices, and higher biodiversity associated with low-intensity land uses, which generate mosaics of wooded and grassy habitats. For example, Swedish semi-natural woodpastures and forest meadows are important examples of relict cultural landscapes in Europe. Paleoecological research has shown a decrease in the diversity of forest taxa within the last centuries as the result of the reduction in grazing and mowing practices and accompanying canopy closure, with detrimental effects on the mixed deciduous forests trees such as lime (linden) and beech. Paleoecological knowledge has been important in the management of these landscapes (Bradshaw and Lindbladh 2005). Similarly, remnants - a function of old grazing practices, such as oak meadows (oak savannas) - are now threatened through the lack of grazing and management (Fig. 1). Dahlström et al. (2008) show that the highest species richness in plants in central Sweden is found in what today is semi-natural grassland subjected to a long continuity of grazing. It is estimated that since 1870, 99.7% of the semi-natural hay meadows have been lost, the few remaining semi-natural hay meadows are now managed mainly for conservation purposes. In this case, long-term data may provide knowledge on management practices that have now been lost.
 |
|
Figure 2: Tuscany landscape. Photo: Martin Falbisoner, Wikimedia Commons. |
In Tuscany, Italy, there has been a 45% decrease of landscape mosaics between 1832-2004, due to the combination of EU regulations in the agricultural and forestry sectors (Agnoletti 2006; Fig. 2). Historical analyses show that meadows and chestnut orchards are being replaced by forest expansion, with an overall loss in landscape heterogeneity, biodiversity and ecosystem services. Pastoral customary practices often combine grazing with fire management to maintain wood pastures, but since the 18th century many state authorities have actively used fire both in agriculture and forestry. Shakesby et al. (2011) showed an increase in wildfire and erosion following rural land abandonment in the Mediterranean region, associated with increased shrub cover and afforestation with flammable species. Paleoecological studies in the Pyrenees have shown that these pastoral landscapes have been systematically managed by fire over millennia (Mazier et al. 2006) and similar results have been shown for the Iberian Peninsula (Gil-Romera et al. 2010). In Scandinavia, the ecological role of fire in both semi-natural and old growth forests is now being realized (Bradshaw and Hannon 2006). In the Finnish boreal forests, dominated by the conifers Scots pine (Pinus sylvestris) and Norway spruce (Picea abies), local slash-and-burn practices were common until the early 20th century. The mixed forests and grazing lands within the boreal forests are now experiencing a degradation and reduction in species diversity as slash-and-burn practices have been discontinued (Myllyntaus et al. 2002). In other parts of Scandinavia, the historical importance of fires in the boreal forests is shown by the presence of fire-adapted species (Bradshaw and Hannon 2006). In all cases listed above, paleoecological knowledge is essential in managing and restoring landscapes, and for the protection and management of rare species through sound management practices (Gillson 2015).
The importance of co-learning
With only 12% of land in protected areas globally, conservation strategies are needed that integrate food production and biodiversity conservation. Intensive agricultural systems provide little wildlife habitat, whereas many land areas under customary management are compatible with biodiversity conservation and provide a wider range of ecosystem services. Cultural landscapes are now an important conservation target, but many customary systems have been eroded due to socio-economic pressures.
Alongside historical studies and stakeholder participation, paleoecological data can help to identify periods of time in which biodiversity and food production co-existed, enabling realistic future scenarios to be envisioned (Gillson 2015). Landscape management needs targets for both the extent and configuration of habitats needed to maintain species, plus political arenas where stakeholders can resolve (the inevitable) conflicts arising from different land-use practices and goals (Angelstam 2006). Rural areas where customary practices are in use today are often either marginal or remote with poorly developed infrastructure, and areas that typically lie very far from the centers of political power where decisions are made. As most biodiversity protection will have to be provided by local initiative and through local practices, we need to better understand and encourage local interest in building and maintaining cultural landscapes. Most importantly, local practitioners, farmers and smallholders or herders must have a place at the negotiating table. Co-learning and co-production of landscape management plans can help to foster ecological and social resilience, as well as nurturing social cohesion and a sense of place, resulting in more effective landscape stewardship.
affiliations
1Department of Archaeology and Ancient History, Uppsala University, and Centre for Environment and Development Studies (Cemus, Uppsala University and Swedish University of Agricultural Sciences), Sweden
2Department of Biological Sciences, University of Cape Town, South Africa
contact
Anneli Ekblom: anneli.ekblomarkeologi.uu.se
references
Agnoletti M (2014) Landscape and Urban Planning 126: 66-73
Bradshaw RHW, Lindbladh M (2005) Ecology 86: 1679-1685
Dahlström A et al. (2008) Env Hist 14: 385-403
EEA (2011) http://www.eea.europa.eu/themes/agriculture
Fischer J et al. (2012) Cons Lett 5: 167-175
Gil-Romera G et al. (2010) Quat Sci Rev 29: 1082-1092
Mazier F et al. (2006) Holocene 16: 91-103
Myllyntaus T et al. (2002) Env Hist 7: 267-302
Peter A. Gell
Long-term data from paleoecology can help identify the “natural ecological character” of wetlands and so inform “limits of acceptable change” and the design of appropriate restoration targets for management that consider the historical range of variability.
While the knowledge of the biodiversity of wetlands is poor, it is understood that, while they cover a small fraction (~6%) of the Earth’s surface, they support a disproportionately rich assemblage of plants and animals. However, freshwater ecosystems are among the most threatened in the world and are suffering biodiversity declines as great, or greater, than terrestrial systems (Dudgeon et al. 2006). Global estimates suggest freshwater vertebrate populations have declined at ~2% per annum since the 1960s through a combination of overexploitation of water resources, pollution, hydrological change, degradation of habitat and invasive species. While these effects vary regionally, it is estimated that 30-90% of the world’s wetlands have been destroyed or strongly modified (Junk et al. 2013). At an international level, the Ramsar Convention is an important framework to protect the world’s remaining significant wetlands.
What is Ramsar?
The Ramsar Convention is a UN program that seeks to conserve the world’s wetlands and their ecosystems. First ratified in Ramsar, Iran, in 1971, it is now represented by 169 countries which have at least one nominated site on the list. The main purpose of the Convention is to mitigate the worldwide loss and degradation of wetlands and this goal is now extended to the wise use of all wetlands within the signatory countries (www.ramsar.org). Today 2243 wetlands are protected under the Convention covering 216,320,717 ha of wetland habitat.
Listing involves the nomination of the site’s boundaries and development of a case that the site satisfies one or more of nine criteria. Mostly the criteria relate to the site’s role in hosting substantial populations of rare or vulnerable fish or bird species or communities. Signatory nations are required to produce a Site Information Sheet and are also encouraged to produce management plans within which they identify the “natural ecological character” of the site. Invariably this description is drawn from the character of the wetland at the time of, or through the period leading up to, listing.
All signatory nations are required to report to the Ramsar Secretariat if a wetland has changed, or is likely to change, from the identified condition. This invokes a pathway of committing to restore the site, offsetting it by nominating an equivalent site, or invoking “national economic interest”. However, despite the Millennium Ecosystem Assessment reporting widespread loss and decline in the world’s wetlands, few nations have reported change in site condition.
The mismatch between the observations of the declining extent and condition of the world’s freshwater systems and the paucity of nation’s reporting that their wetland has changed, or is likely to change in the future, could be interpreted as all declines occur outside the network of listed sites. This would confer considerable merit to the Ramsar Convention in mitigating the global decline in condition. However, what is more likely is that nations have been either unable to detect change, or have been reluctant to report because of the challenges of restoration, and how it may reflect their record of environmental management. In order to drive better reporting of the condition of a wetland relative to this baseline, the Convention has requested that nations identify the limits of acceptable change.
 |
|
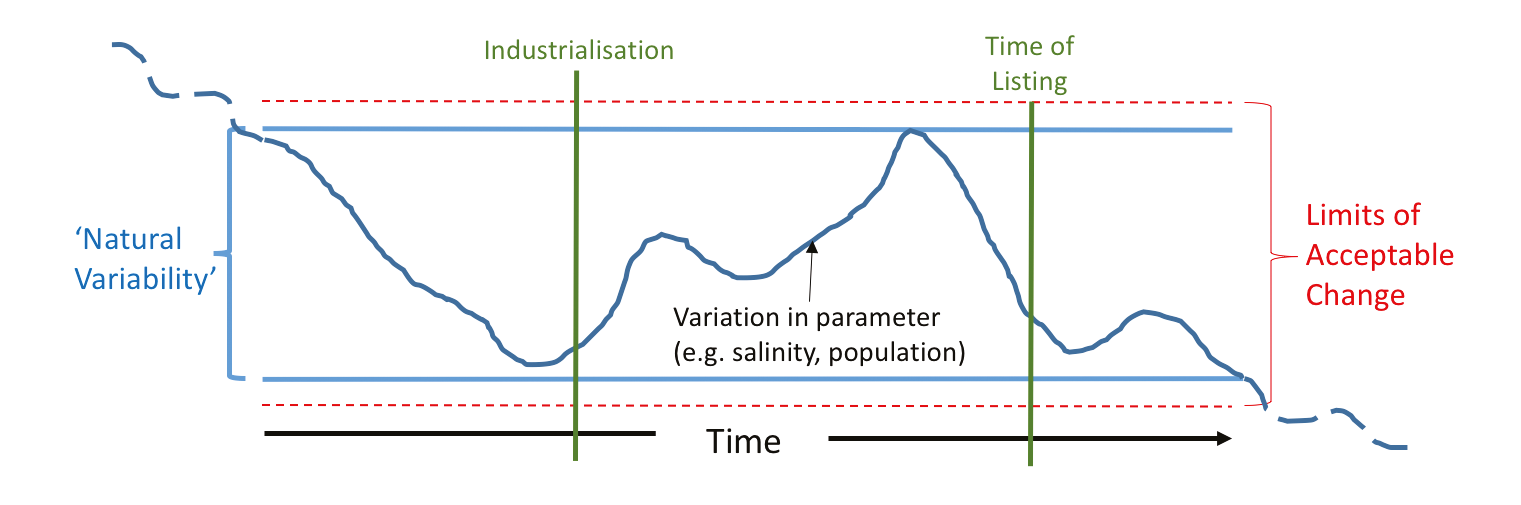
Figure 1: Theoretical approach to developing “Limits of Acceptable Change” within Ramsar wetlands (adapted from Phillips 2006). In principle, the limits of acceptable change are set just outside the margins of the range of condition as recorded from history. The dashed lines illustrate that longer cycles of change may challenge even this record of natural variability. More realistically, there is little continuous monitoring data that extends far beyond the time of listing to realistically assess variation in any parameter, and so paleoecological records are essential in defining limits. |
Means of determining the limits of acceptable change have been proposed based on the historical variability in a measured condition in the past (Fig. 1). However, the monitoring of wetland condition and populations of aquatic flora and fauna has a short history, even among developed nations with a strong tradition of aquatic-system research. Also, the influence of low frequency cycles of change question the representativeness of records of variability that may even span several decades. Paleoecological approaches can, and have, extended this knowledge of variability and so play an important role in the understanding of wetland change over time.
Paleoecology and Ramsar wetlands
There have been many records of paleoecological research on Ramsar wetlands, although it is possible that not all researchers were aware of the status of the site at the time of listing, relative to its historical past. Evidence from the Coorong, Lower Lakes and Murray Mouth site in Australia revealed that the site description at the time of listing in 1985 reflected a condition that had changed in the 1950s (Reeves et al. 2015) and was unrepresentative of what would be considered the natural long-term baseline (Gell 2017). Ramsar sites along the Nile Delta have also been shown to have been substantially impacted in recent decades and that the present condition is a departure from the long-term baseline (Flower et al. 2001).
Paleoecological research can also provide direct evidence on the status and breeding success of significant waterbird populations. Loch Ruthven near Inverness in Scotland was listed as a Ramsar site in 1996 mainly by way of the important population of breeding Slavonian Grebe it hosts. Brooks et al. (2012) used a sediment record to relate the annual recruitment success of Slavonian Grebe, based on historical surveys, to the availability of chironomid prey for the hatchlings, as recorded in the lake sediments. The increased nutrient load is linked to larger populations of this key food item for the Grebe chicks leading to enhanced recruitment and securing one of the main criteria of the loch’s listing.
PAGES has a long history of documenting the impact of people and climate on wetlands through themes, projects and working groups e.g. LIMPACS and Aquatic Transitions. Many records attest to considerable condition change both before and since the Convention began. So, from a paleoecological perspective, it is clear that long-term records have the capacity to provide better understanding of present condition in the context of variability, including the impacts of low-frequency climate cycles, as well as providing records of shifting baselines due to human impact. Acknowledging this, PAGES and the Ramsar Secretariat hosted a workshop in Queenscliff, Australia, in late 2013 to examine the opportunities that may exist, for the Convention and for paleoecological researchers, to bring a long-term perspective to the identification of character and the setting of acceptable limits to change based on knowledge of long-term variability. The proceedings of this workshop are published in a special issue of Marine and Freshwater Research (Gell and Finlayson 2016), including an editorial from the previous Ramsar Convention Secretary General (Davidson 2016) declaring the benefits that paleoecology can bring to more realistic assessments of the world’s wetlands of international significance.
Shifting baselines and limits of acceptable change
While it remains challenging for nations to identify how current wetland conditions have deviated from long-term trends, if only a recent state is used as a baseline the failure to consider pre-listing state allows for all of the previous human impact to be excused, potentially perpetuating mismanagement in a degraded state (Finlayson et al. 2016). The Millennium Ecosystem Assessment documented widespread reduction in the extent and condition of freshwater wetlands worldwide, so it is implicit that time-of-listing condition is unlikely to reflect the range of wetland conditions before impact by industrialized society. Without long-term data, nations might continue to manage for a derived, and perhaps less productive, wetland than previously existed.
 |
|
Figure 2: Proposed pathway to guide action following the identification of change to a Ramsar listed wetland. Paleoecological approaches can assist in assessing if the driver of change is within or outside the jurisdiction of the signatory party, whether it is within or outside limits of acceptable change (LAC) and whether it is in response to global climate change or a cross border effect. Nations are required to restore or offset when the change is outside long-term variability and if the driver of change is the responsibility of the signatory nation. ECD: Ecological character descriptions. Photo: The Coorong Ramsar site, Australia; Peter Gell. |
Conversely, armed with a better understanding of variability and defined limits of acceptable change, a nation is better equipped to decide when and how to act, and when to report change as being outside, or within, past historical range. Further, it is unreasonable if a nation is held to account for the change in the state of a wetland if the driver of that change is due to the actions of a neighbor (e.g. upstream), or is global (e.g. climate change) in scale. By partitioning human impact from climate, and proximal and distal causes of change, paleoecology can assist with an appropriate pathway for the Ramsar Secretariat to assess the obligation of nations to act (Fig 2).
affiliations
Water Research Network, Federation University Australia, Ballarat, Australia
contact
Peter A. Gell: p.gellfederation.edu.au
references
Brooks SJ et al. (2012) J Paleolimn 47(4): 631-644
Davidson NC (2016) Mar Fresh Res 67: 685-686
Dudgeon D et al. (2006) Biol Rev 81: 163-182
Finlayson CM et al. (2016) Mar Fresh Res 67: 687-694
Flower RJ et al. (2001) Aq Ecol 35: 369-388
Gell PA, Finlayson CM (2016) Mar Fresh Res 67(6)
Junk WJ et al. (2013) Aq Sci 75: 151-167
Phillips B (2006) Critique of the Framework for describing the ecological character of Ramsar Wetlands. Mainstream Environmental Consulting Pty Ltd
Bronwen S. Whitney1 and Macarena L. Cárdenas2
Impressive archaeological sites, often underlying dense forest, show evidence of past human impact within “pristine” ecosystems. Here we present four examples that highlight the challenges faced by paleoecologists in determining the potential legacies of human impact on these ecosystems.
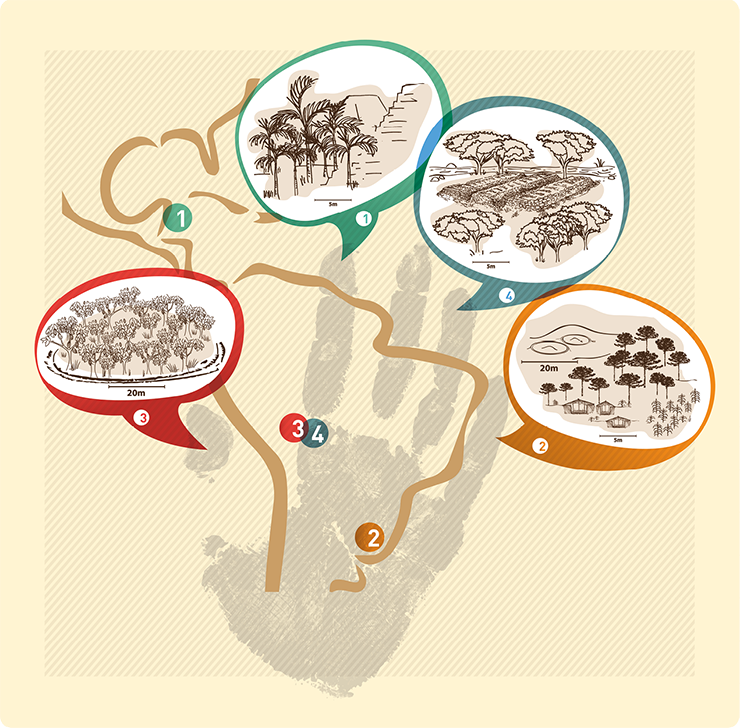 |
|
Figure 1: Locations of selected case studies in Latin America and illustrations of the land-use type and key plant species for each scenario. |
Across Latin America, the uncovering of impressive archaeological sites, often underlying dense forest, shows evidence of past human impact within what are often viewed as pristine ecosystems (Bush and Silman 2007; Willis et al. 2004). Anthropogenic impacts were spatially heterogeneous and it is often argued that modern floristic composition in areas of past human occupation espouses a legacy effect in terms of an abundance of economically useful species. Many argue that we need to shift our perceptions of what is “natural” in modern tropical ecosystems and to consider indigenous knowledge and agencies in management and conservation strategies (Clement et al. 2015). This new perspective challenges us to take into account how vegetation diversity and composition in human-impacted areas have responded and changed according to different prehistoric land-use strategies. Most studies of legacy effects rely on modern floristic surveys (Ford and Nigh 2009) that compare composition in areas inferred to have experienced high and low impact in the past. A key limitation of the modern comparative approach is the lack of temporal dimension to the studies. Paleoecology can offer unique insights into legacy impacts on forest composition and biodiversity because it affords a longer temporal perspective than offered by comparative floristic studies and conventional ecology, and in most cases, the studies pre-date complex societies, so they can provide an ecological baseline. Here we review four case studies from the tropical and subtropical Americas (Fig. 1) that demonstrate how paleoecology is attempting to address the question of legacy effects on ecosystems that experienced varied pre-historic land-use strategies.
Ancient Maya palm cultivation
 |
|
Figure 2: Selected data from the case studies showing the impact and legacy of Pre-Columbian land use on tropical ecosystems. Data are presented as percent abundance of all pollen, except in the case of large, rare pollen grains, such as Inga and maize, where individual counts are shown. (1) High palm abundance during the Classic Maya period in Lamanai (Rushton et al. 2012); (2) Very high Araucaria pollen coincides with the establishment of the Jê culture and increase in disturbance taxa (Mimosa), suggesting anthropogenic changes in the forest structure; (3) Forest expanded around the time of the establishment of “chocolate forest islands”, but without evidence of Theobroma pollen in the record, we cannot determine if the high abundance of these trees occurred due to past cultivation; (4) Consistent Inga pollen after the shift from crop cultivation suggests this culture might have promoted the growth of Inga as a food source. |
Questions over the persistence of past human impacts are no better illustrated than among studies of the seasonal forests of the ancient Maya lowlands. The issue of legacy impacts in Maya forest (Lambert and Arnason 1982) initially focused on Breadnut (Brosimum alicastrum, Moraceae), an economically important tree known locally as ramón, that occurs in high proportions in forests surrounding temple ruins, but their high abundance was linked to calcareous soils created by crumbling temple ruins, rather than direct human management. More recently, convincing evidence has emerged of higher abundance of economically important plant species, such as palms, in ancient forest gardens in Belize compared to high bush forest sites (Ross 2011). Few paleoecological studies have addressed legacy impacts on forest composition and biodiversity in the Maya lowlands, with most concerning forest clearance and agriculture at the height of the Classic Maya period. However, a detailed pollen study of Lamanai, Belize, showed clear evidence of palm monoculture (Fig. 2) at the height of the Classic period (Rushton et al. 2012). Palms are vitally important plants that have a multitude of uses, including thatch, nuts, oils, and dyes. The hyperdominant occurrence of several economically important palm species across the Neotropics have been presented as evidence of their wide spread use in prehistory. However, the high palm abundance in this pollen record did not persist into the modern era. As a consequence of the reduced human manipulation of the landscape, the palm plantation rapidly reverted to a more diverse seasonal broadleaf forest, suggesting that human intervention is required for monodominant palm stands to persist.
The legacy of human impact in Araucaria forest
Among the ridges of the subtropical southern Brazilian highlands, the living fossil Araucaria angustifolia tree dominates the landscapes that were once home to the complex Jê society that emerged at around 2,000 yr BP. Araucaria or the “monkey puzzle tree”, provided to the Jê culture not only wood and shelter for their villages, but also one of their staple foods, the pinhão, a highly nutritious and calorie-dense nut used in their daily diet as well as in rituals and ceremonies (Iriarte et al. 2013). Paleoecological studies show that Araucaria trees arrived in the south of Brazil from about 6,000 yr BP, and expanded following the climatic trend of increased precipitation through the Holocene (Behling et al. 2004). Meanwhile, the paleoecological records within Jê archaeological context shows unexpected patterns in the diversity of this forest and the abundance of Araucaria pollen if climate was the sole driving factor. The ongoing studies show that the more intense and permanent occupation of this region was associated with a regional expansion of the Araucaria forest at the same time that this culture modified the landscape at a local scale (Behling et al. 2004; Iriarte et al. 2013; Iriarte and Behling 2007). The varied proportion of Araucaria forest at human-impacted sites could reflect the legacy of pre-Columbians having facilitated the spread of this economically valuable tree species (Iriarte and Behling 2007). It is suspected that the heterogeneous local human land modification across the Jê landscape affected the diversity of “natural” vegetation (Iriarte and Behling 2007), suggesting past land-use legacies have influenced the modern vegetation.
Chocolate forest islands in SW Amazonia
Palm agroforestry by the ancient Maya may not be the only example of monodominant cultivation, as shown in the case of the “chocolate forest islands” in the Llanos de Moxos of southwestern Amazonia. The Llanos de Moxos is an Amazonian sub-basin that has been inhabited as early as 10,000 yr BP, with maize crop production beginning ca. 6,500 yr BP (Brugger et al. 2016). The basin contains impermeable seasonally-flooded soils overlain by savannas; although much of the region is not hospitable for forest growth, there are forest islands on well-drained soils of outcrops of the PreCambrian Shield located within savanna. One such forest island, where a ring-ditch traces the entire periphery of the forest island, shows clear evidence of past human manipulation of the landscape (Carson et al. 2016). Intriguingly, dense stands of cacao (Theobroma cacao) occupy a large proportion of the island, which are thought to originate from either 17th century Jesuit missionaries or Pre-Columbian agroforestry. Due to the highly-specialized pollination mechanism, however, Theobroma pollen was undetectable in the pollen record, even with counts of > 1000 grains, so the origin of the chocolate plantation could not be dated. What paleoecology could reveal, however, is that the dense broad-leaf forests of southwestern Amazonia became established in the late Holocene (3,000 yr BP) due to increasing monsoon strength, long after anthropogenic impacts had begun in this region. Given that the rainforest in this southwestern corner of Amazonia does not pre-date anthropogenic influence, composition of forest in areas adjacent to human habitation might have been influenced by people from its inception, thus the ecological baselines could be anthropogenic.
Cultivation of the ice-cream bean
Also among the lowland savannas of southwestern Amazonia, extensive Pre-Columbian raised fields show that people once cultivated these poor savanna soils where cattle ranching now predominates (Lombardo et al. 2011). Paleoecological analyses show that intensive land use began before 1700 yr BP, with burning and clearance of savannas and gallery forests, and raised fields were built for maize cultivation (Whitney et al. 2014). A shift towards less intensive land use incorporating sweet potato cultivation occurred around 700 yr BP, and accompanying this change, Inga species colonized the riparian zones. Inga edulis is a common food resource (the ice-cream bean) and also an ecological pioneer in the establishment of secondary tropical forest. Pioneer tree species tend to out-shade themselves in the first decades following forest regeneration (Guariguata and Ostertag 2001), so its centuries-long presence in the paleoecological record points to human management of the regenerating gallery forest that stopped forest succession towards a more diverse floristic composition. The mixed strategy of agriculture and agroforestry continued until 1800 AD, around the time land use switched to cattle ranching, suggesting that continued management was required for Inga species to persist.
This short review of Pre-Columbian impact on ecosystem composition and biodiversity shows variable evidence for legacies of past land use. The intensity and type of past management, combined with ecosystem type, might be key to controlling whether or not anthropogenic modifications create long-term changes to ecosystem composition and biodiversity. Despite having made significant inroads in redefining what is natural in tropical ecosystems, the complexity illustrated by these studies highlights the challenges ahead for paleoecology in disentangling the natural and human determinants of composition and biodiversity in tropical ecosystems.
affiliations
1Department of Geography and Environmental Sciences, Northumbria University, Newcastle-Upon-Tyne, UK
2Department of Geography and Environmental Science, University of Reading, UK
contact
Bronwen Whitney: bronwen.whitneynorthumbria.ac.uk
references
Behling H et al. (2004) Palaeogeog Palaeoclimat Palaeoecol 203: 277-297
Brugger SO et al. (2016) Quat Sci Rev 132: 114-128
Bush MB, Silman MR (2007) Front Ecol Environ 5: 457-465
Carson JF et al. (2016) J Quat Sci 31: 337-347
Clement CR et al. (2015) Proc R Soc B 282: 20150813
Ford A, Nigh R (2009) J Ethnobiol 29: 213-236
Guariguata MR, Ostertag R (2001) Forest Ecol Manag 148: 185-206
Iriarte J, Behling H (2007) Environ Archaeol 12: 115-127
Iriarte J et al. (2013) J Anthropol Archaeol 32: 74-96
Lambert JDH, Arnason JT (1982) Science 216: 298-299
Lombardo U et al. (2011) J Archaeol Sci 38: 502-512
Ross NJ (2011) Ecol Appl 21: 75-84
Rushton EA et al. (2012) Holocene 23: 485-493
Valentí Rull1, T. Vegas-Vilarrúbia2 and E. Montoya1
Late Pleistocene and Holocene paleoecological reconstructions from the neotropical Gran Sabana provide objective criteria to assess realistic and feasible restoration targets, and to appraise the potential effects of global warming on the landscapes and ecosystems.
A significant contribution of paleoecology to conservation and restoration practices is its ability to provide evidence of past ecological conditions, thereby permitting the identification of baselines, to which it would be desirable to return. Choosing the most desirable target is the first step in restoration planning, and a frequent option is to choose to restore the ecological conditions that existed prior to human disturbance (Willis et al. 2010). Another relevant conservation output that may be derived from paleoecological records is the response of ecological communities to past climate change as a model for anticipating potential ecological responses to future climate change (Vegas-Vilarrúbia et al. 2011). Here, we use these approaches to analyze the case of the Gran Sabana region, located in northern South America.
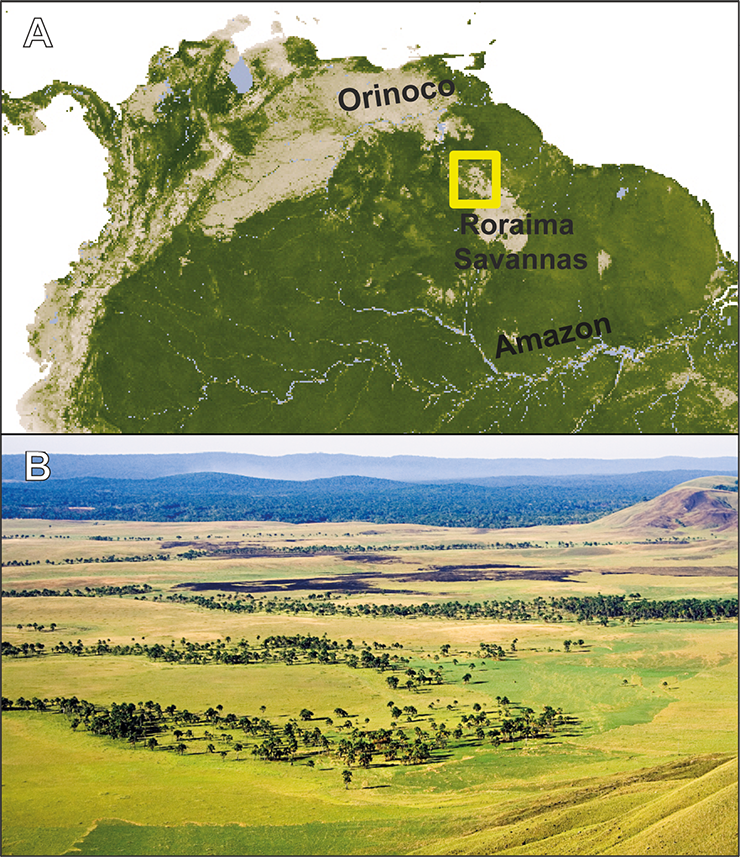 |
|
Figure 1: The Gran Sabana. (A) Map of northern South America showing the Roraima Savannas and the Gran Sabana (yellow box), between the Orinoco and Amazon basins. (B) Typical landscape of the Gran Sabana savannas and Mauritia palm swamps with dark patches corresponding to recent fires. The Orinoco-Amazon rainforests are in the background. Photo: V. Rull. |
The Gran Sabana is part of a huge savanna island within the Amazon-Orinoco rainforests (Fig. 1). Most of the Gran Sabana is covered by treeless savannas, with forest-savanna mosaics at the edges and gallery forests along rivers. A special type of gallery forest is the emblematic palm swamp of Mauritia flexuosa, locally known as “morichal” (Rull and Montoya 2014). The Gran Sabana is part of the Canaima National Park, in the headwaters of the Caroní River, a tributary of the Orinoco that is exploited for hydroelectric power. The savannas are nowadays in expansion to the detriment of forests, mainly due to burning by indigenous people, sparking a debate between defenders and detractors of these practices. Defenders argue that fire management is a fundamental part of indigenous traditions and should be preserved, while critics believe that fires have been responsible for the disappearance of the hypothetical ancient forests that covered the Gran Sabana before the onset of fire practices. This debate is relevant for conservation, as the detractors of indigenous practices argue that forest clearance should cease, whereas the indigenous defenders believe that fires are useful for land management and should be included in conservation planning (Sletto and Rodríguez 2013).
Paleoecology and restoration targets
The Gran Sabana paleoecological records extend back to the Younger Dryas, when it was covered by a mosaic of forests, shrublands and savanna patches. The vegetation cover was mostly ligneous but a continuous forest cover was not observed (Montoya et al. 2011a; Rull et al. 2013). The extensive savannization process started in the Younger Dryas-Holocene transition and culminated by ca. 10 cal ka BP (Fig. 2). Fires, possibly of anthropogenic origin, and the Early Holocene Warming seem to have been involved in this savannization trend, which was relatively rapid and hitherto irreversible (Rull et al. 2015a). During most of the Holocene, fires remained at low rates and the Gran Sabana was dominated by treeless savanna with gallery forests and without Mauritia palm swamps. Gallery forests experienced a significant increase during the middle Holocene (ca. 7 to 5 cal ka BP), likely due to a general increase in the available moisture (Montoya et al. 2011b; Rull et al. 2016). Mauritia flexuosa arrived to the Gran Sabana by 2 cal ka BP, coinciding with a significant exacerbation of fire, considered to be the onset of the present-like indigenous fire management practices, and the establishment of modern Gran Sabana landscapes (Montoya and Rull 2011; Montoya et al. 2011c; Fig. 2). During the last 2000 years, Mauritia swamps have experienced a remarkable expansion because they are not burnt by indigenous people, who use this palm for multiple purposes (Rull and Montoya 2014).
 |
|
Figure 2: Synthesis of the main paleoecological trends during the last 13,000 years. The ecological states i, ii and iii are described in the text. Modified from Rull et al. (2016). |
In summary, we have identified three general ecological states since the Late Glacial: (i) a forest-shrubland-savanna mosaic dominated by ligneous communities during the Younger Dryas, (ii) extended treeless savannas with gallery forests during most of the Holocene, and (iii) treeless savannas with gallery forests and Mauritia palm swamps during the last 2000 years. It is possible that the number of ecological states will increase as further studies on glacial and former times are conducted. Two other potential states not observed in the available past ecological records are: (iv) treeless savannas totally devoid of forests and (v) continuous forest cover without savannas. On the basis of paleoecological studies, both of these scenarios would be unrealistic as a conservation target.
Restoration proposals
Using the criterion of pre-human conditions (target i) would be problematic, because Younger Dryas climatic conditions differed from those of today, and because this would be in conflict with present-day indigenous practices. Restoring the prevalent Holocene conditions (target ii) would necessitate the removal of the emblematic Mauritia palm swamps, which would be a very unpopular proposal, involving the destruction of the present-day Gran Sabana landscape that is generally regarded as “natural”. Moreover, it would be difficult to develop such actions in a national park created on the basis of current landscape features. In addition, Mauritia flexuosa is a key resource for indigenous people over the entire Neotropics (Rull and Montoya 2014), and its removal would be viewed as cultural aggression. The third option (target iii) - maintaining the Gran Sabana as it is at present - implicitly accepts the continued use of fire by the indigenous people, a condition that would not be acceptable to the defenders of the assumed continuity of former hypothetical rainforests. Because it does not require any special action, this option would be the least expensive in terms of budget and effort; however, its likely consequence will be the continued expansion of savannas to the detriment of rainforests. An alternative solution, at least in theory, would be the joint recreation of present and former ecological states (i, ii and iii) in different parts of the Gran Sabana. This option has been called conservation quasi in situ or inter situ (Burney and Burney 2007; Volis and Blecher 2010); its main drawbacks are that large investments are required to create and maintain such structures and that it would imply intervention into indigenous land-use practices, which is very problematic under the current socio-political circumstances (Rull et al. 2016).
Responses to global warming
The Younger Dryas-Early Holocene warming has been proposed as a past analogue of future global warming in the Neotropics (Rull et al. 2015b). According to the latest IPCC report, by 2100 the Gran Sabana region will experience an average temperature increase of 2-3°C, whereas precipitation will remain at similar-to-present values, with a range of variation of 20% (IPCC 2013). This scenario is similar to the Younger Dryas-Early Holocene warming transition, when treeless savannas already dominated the Gran Sabana landscape and the incoming of wetter climates was not sufficient for forest recovery, likely due to fire continuity. As stated earlier, this savannization process was irreversible until today. Using this model, it can be predicted that continuation of present-day fire practices will likely result in the irreversible expansion of savannas and a concomitant reduction of forests, similar to that observed historically. Therefore, fire control measures should be agreed upon with indigenous people if the remaining Gran Sabana forests are to be preserved in their present state (target iii). If a landscape of treeless savanna devoid of forests and spiked with morichales is the desired conservation target (scenario iv), then no action is required. Again, the recovery of the pre-human Younger Dryas landscape (target i) seems unattainable under the IPCC predictions, using the past analogue scenario depicted here. For similar reasons, total reforestation of the Gran Sabana (scenario v) would be unworkable.
Conservation policy
All these considerations and others that have emerged from paleoecological studies should be evaluated by the stakeholders and policy-makers as a basis for developing the best conservation strategy. The role of paleoecologists is to provide the necessary information on aspects of temporal ecological dynamics that may be pertinent to conservation and possible restoration targets. Paleoecological information should be part of these deliberations; otherwise, conservation targets will be unrealistic and landscape development under the action of environmental drivers will be unpredictable.
affiliations
1Laboratory of Paleoecology, Institute of Earth Sciences Jaume Almera (ICTJA-CSIC), Barcelona, Spain
2Department of Evolutionary Biology, Ecology and Environmental Sciences, University of Barcelona, Spain
contact
Valentí Rull: vrullictja.csic.es
references
Burney DA, Burney LP (2007) Front Ecol Environ 5: 483-490
IPCC (2013) Climate Change 2013. The Physical Science Basis, Cambridge University Press
Montoya E, Rull V (2011) Quat Sci Rev 30: 3430-3444
Montoya E et al. (2011a) J Quat Sci 26: 207-218
Montoya et al. (2011b) Palaeogeogr Palaeoclimatol Palaeoecol 310: 413-426
Montoya et al. (2011c) Quat Res 76: 335-344
Rull V, Montoya E (2014) Quat Sci Rev 99: 17-33
Rull V et al. (2013) Persp Plant Ecol Evol Syst 15: 338-359
Rull V et al. (2015a) Quat Sci Rev 122: 158-165
Rull V et al. (2015b) Quat Sci Rev 115: 28-38
Rull V et al. (2016) Holocene 26: 1162-1167
Sletto B, Rodríguez I (2013) J Environ Manag 115: 155-166
Vegas-Vilarrúbia T et al. (2011) Quat Sci Rev 30: 2361-2388
Rob Marchant, C. Courtney-Mustaphi and E. Githumbi
Under current rapid environmental change there are numerous challenges for people and wildlife. Paleoecological records from southern Kenya are synthesized to provide understanding on how abiotic and biotic elements intertwine and can provide a foundation to inform decisions for environmental management.
Abiotic and biotic elements intertwine over multiple spatio-temporal scales to determine the present-day composition, structure and distribution of ecosystems. Rapid compositional and distributional changes, particularly during the past few hundred years, are attributed to anthropogenic modifications that are superimposed on long-term climatic and landscape changes (Dearing et al. 2010). It is crucial to have a paleo perspective to disentangle interactions of how the environment, ecosystems, animals and people have combined to influence current ecosystem states, and to learn how lessons from the past can be used to predict and constrain possible future trajectories (Marchant and Lane 2014).
The savanna ecosystems of East Africa are characterized by relatively rapid population growth, changing political, economic, social and cultural contexts, and an agricultural system that is increasingly focused on relatively recently adopted crops, such as maize and market gardening, for regional and international export. This expansion and intensification of agriculture across East African savannas induces particularly rapid land-use and land-cover change. East African savannas also support large human and herbivore populations and have become frontiers for water-intensive agricultural expansion on communal lands formerly dedicated to livestock grazing. Balancing economic development while maintaining ecosystem integrity and the globally important protected area creates policy challenges, which are further exacerbated by climate change. The traditionally reliable dry and rainy seasons have been increasingly disrupted and the unpredictability of the rainy season has become the norm for many savanna communities. Government policies that have encouraged formerly mobile herders to adopt more settled ways of life for both positive (health, education) and negative (land grab, containment) reasons, combined with land fragmentation and degradation, hamper the adaptive ability of communities to respond to inherent climatic variability, for example through transhumance pastoralism. Increasingly intensive land uses, often focused around Protected Area boundaries (Pfeifer et al. 2012), similarly pose threats and challenges for ecosystems, conservation and livelihoods. While there have been numerous calls to address such challenges, current intervention strategies have had limited success, as evidenced by continued encroachment, human-wildlife conflict and the rise of poaching and illegal hunting, along with unabated rapid agricultural development and associated pressure on water resources. These changes are leading to (wildlife) habitat degradation, increased fragmentation of populations, decreased resilience, and biodiversity loss across much of the East African savannas.
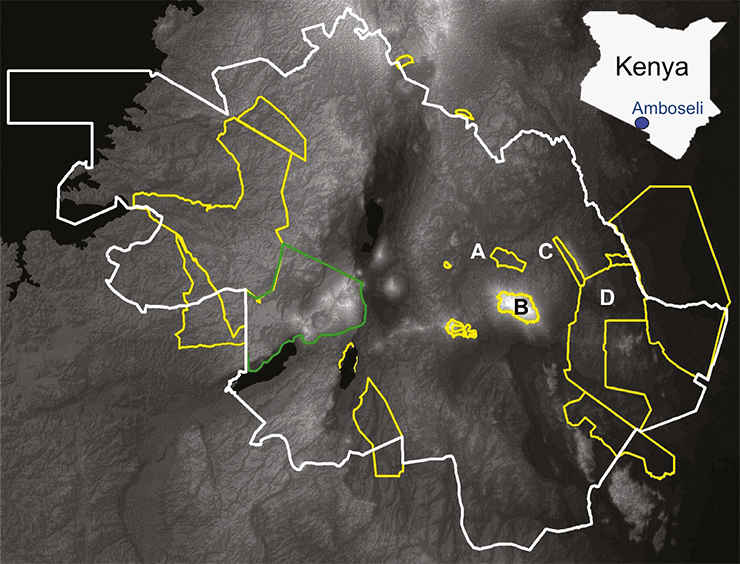 |
|
Figure 1: Map of the Kenya-Tanzania Borderlands within the white lines showing the outline locations of the National Parks (yellow) and the Ngorogoro Conservation Area (green). The protected areas (yellow) that surround the Amboseli National Park (A) are Kilimanjaro (B), the Chyulu Hills (C) and Tsavo National Parks (D). |
One area where these transformations have been acutely felt is within the Amboseli landscape, an area that is somewhat symptomatic of the wider Kenya Tanzanian borderland area (Fig. 1). The Amboseli landscape supports the highest wildlife densities in southern Kenya that migrates across the Kenyan and Tanzanian border. Within the Amboseli landscape there are several perennial wetlands distributed across the predominantly semi-arid landscape; these groundwater-fed wetlands, recharged from orographic precipitation falling on Mount Kilimanjaro (Fig. 2), provide an important source of water and wildlife refuge, and form a series of “stepping stones” for animal migrations between Amboseli National Park and the neighboring Tsavo and Chyulu Hills National Parks (Fig. 1). Unlike seasonally rain-fed wetlands, which are inundated during the wet season and dry out during the dry season, the Amboseli wetlands sustain perennially high local water-tables that enable peat accumulation through drought periods and hence form a paleoecological archive. Reconstruction of the vegetation, climatic and fire histories has been carried out on five of these swamps through pollen, non-pollen palynomorph and macroscopic charcoal analysis, which combine to produce a landscape scale understanding of ecosystem transitions and unpick the drivers behind these shifts. From the mid Holocene, East Africa shifted towards a drier environment characterized by reduced precipitation, increased evaporation, and/or an extension or intensification of the dry season. This period of regional aridity is clearly marked by a peak in aeolian dust deposition within the Kilimanjaro glacier dated to around 4000 cal yr BP (Thompson et al. 2002). Under these relatively arid conditions, the Amboseli ecosystems were dominated by semi-arid taxa (Acacia, Aloe, Amaranthaceae-Chenopodiaceae, Asteraceae and Capparis). Low charcoal concentrations due to low fuel availability show further evidence of the sparsely vegetated nature of the Amboseli landscape. Between ~2000 and 800 cal yr BP, there is an increase in pollen diversity as the savanna ecosystem became more dominated by woodland taxa (Acalypha, Balanites, Commiphora, Cordia and Salvadora), and aquatic taxa (Cyperaceae and Nymphaea) became locally more abundant. Increased abundance of wind-blown Afromontane forest taxa (Celtis, Juniperus and Olea), most likely derived from the adjacent slopes of Mount Kilimanjaro and the Chyulu Hills, further suggests a period of increased moisture that was concomitant with increased local biomass burning. After c. 800 cal yr BP, a drier environment returns with the increased presence of Amaranthaceae and Poaceae and decreased levels of mesic tree taxa such as Syzygium (Rucina et al. 2009), again with a regional expression of lower lake levels (Lamb et al. 2003; Westerberg et al. 2010).
 |
|
Figure 2: Different perspectives on Amboseli National Park. (A) The area is dominated by Mt Kilimanjaro; this is also the “water tower” that feeds the swamps with water to allow accumulation of sediments and the past ecosystem histories to be preserved. (B) Herbivores, particularly elephants, are crucially important ecosystem engineers across the landscape: the exclosure experiment clearly shows that with ca. seven years respite, a grazing lawn transitions into tree-dominated savanna (to the right of the fence). The wholescale removal of elephants from the East African landscape, particularly over the past four centuries, will have had a transformative impact on savanna structure and composition. (C and D) Current land-use transformations are similarly having a large impact on the swamps. Intact wetlands within the National Park boundaries (C) provide a grazing resource, particularly vital during dry seasons that are becoming increasingly severe and prolonged. They are a stark contrast to wetlands just outside the protected area that are drained and converted to agricultural production (D). |
Increased presence of Acacia, Amaranth-aceae/Chenopdiaceae, Balanites and Poaceae in the uppermost samples dating to the last ca. 300 years could be indicative of a drier environment, but could also be a consequence of decreased elephant populations which have a major influence on vegetation composition, distribution and structure (Håkansson 2004; Fig. 2). Regional records, for example from Mt. Shengena located to the south of Amboseli in the Eastern Arc Mountains, reflect increased anthropogenic use of the landscape through the appearance and increased abundance of cultivated taxa such as Maize (Zea mays), Castor oil bean (Ricinus) and Pine (Pinus) to meet the needs of a growing and mobile population (Finch et al. 2016). Moreover, caravan trade resulted in the decimation of East African elephant populations (Håkansson 2004) that clearly would have been transformative for forest cover (Fig. 2). Anthropogenic modifications to the ecosystem have intensified dramatically in recent decades. Wetland areas were converted into agricultural production (shambas and commercial farming) as marked by the influx of crops and ruderal taxa (Fig. 2). Pastoralist populations with historical formal and informal land tenure have recently reorganized into more sedentary group ranches where some groups and individuals maintain pastoral livelihoods on an increasingly fragmented landscape. People have also migrated into the region and the increased population has placed additional demands on the Amboseli wetlands, clearly affecting socio-ecological resilience and wider ecosystem functioning of the landscape through the past ca. 4000 years.
It is clear from the Amboseli case study that long-term paleo perspectives provide understanding of the interactions between environment, ecosystems, animals and people, and as such provide a foundation to inform sustainable environmental management decisions (Gillson and Marchant 2014). Understanding how best to manage ecological systems, to protect biodiversity and benefit the human communities that rely on them, requires an interdisciplinary approach; this management is strengthened where paleo perspectives combine with social sciences and humanities to examine complex socio-ecological systems. To strengthen adaptive capacity, and promote resilience, it is crucial to understand how past and present socio-ecological systems are interlinked. Such insights are crucial to inform planning and practice, and inform longer-term planning frameworks such as the UN Sustainable Development Goals.
affiliations
York Institute for Tropical Ecosystems, University of York, UK
contact
Rob Marchant: robert.marchantyork.ac.uk
references
Dearing JA et al. (2010) Ecol Soc 15: 21
Finch J et al. (2016) Holocene 27: 796-810
Gillson L, Marchant R (2014) Trends Ecol Evolut 29: 317-325
Håkansson N (2004) Hum Ecol 32: 561-591
Lamb H et al. (2003) Holocene 13: 285-292
Marchant R, Lane P (2014) J Arch Sci 51: 12-21
Pfeifer M et al. (2012) PloS One 7: e39337
Rucina SM et al. (2010) Holocene 20: 667-677
Daniele Colombaroli1,2,3, C. Whitlock4, W. Tinner1 and M. Conedera5
Projected climate and land-use changes challenge ecosystem management and conservation strategies to maintain biodiversity. Information on past vegetation and fire responses to human activity and climate variability helps identify which conservation targets (natural or cultural) best meet landscape management goals.
Ecological mitigation and restoration efforts aimed at reducing the effects of excessive land use are currently under revision in light of climate projections that suggest significant changes in vegetation structure and composition in the coming decades (Diffenbaugh and Field 2013). Given the magnitude of projected change, future ecosystem dynamics are often difficult to infer on the basis of short-term observations alone (Willis and Birks 2006) and limited time perspectives may result in poor conservation decisions with unexpected consequences under new climate conditions (Gillson et al. 2013; Williams and Jackson 2007). The information offered by historical ecology and paleoecology, spanning centuries to millennia, reveals a range of ecosystem processes occurring at different spatial and temporal scales, as well as the extent to which present vegetation has been altered by past human activities. Understanding past ecosystem dynamics, and particularly the role of climate and humans, offers a baseline for interpreting current landscape conditions and helps set goals for conservation and restoration. Conservation actions will vary depending on (1) where the current landscape falls along a land-use gradient from pristine to intensely altered; (2) the motivation (or cultural values) to achieve more natural, or maintain, cultural landscapes; and (3) a cost-benefit analysis of pursuing particular strategies. We provide two examples of how paleoecology can help identify appropriate restoration targets and discuss the role of past environmental reconstructions for restoration, forest management and biodiversity conservation.
Maintaining nearly pristine landscapes
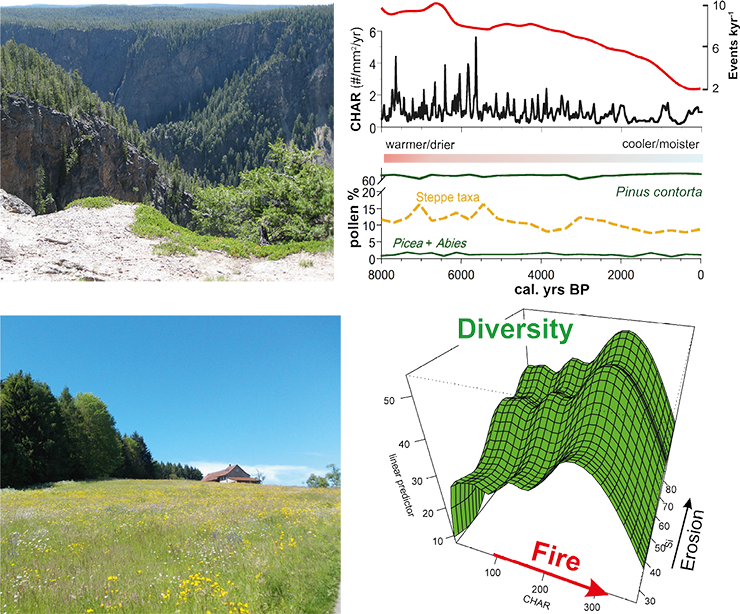 |
|
Figure 1: Top. Much of Yellowstone National Park is underlain by infertile rhyolite substrates that support lodgepole pine (Pinus contorta), a species that is also highly adapted to fire. Pollen and charcoal data suggest that lodgepole pine has maintained dominance in these geologic settings throughout the Holocene despite dramatic climate-driven changes in fire activity (Millspaugh et al. 2000; right panel, photo by CW). Bottom. Highly diverse meadows in mountain ecosystems offer important provisioning and cultural services (Broye Valley near Moudon, Switzerland, photo by DC). On the right: long-term records from the Valais (Central Alps) show variations in biodiversity (based on pollen data) as a function of fire (based on charcoal) and erosion (based on magnetic susceptibility). Plant diversity is maximized at intermediate levels of fire disturbances, in accordance with ecological theory. After Colombaroli et al. (2013). CHAR refers to charcoal accumulation rates, a measure of fire activity. |
Protected areas, like some of the large US national parks, are good examples of nearly pristine ecosystems, in that they support most of the species and ecological processes that existed prior to Euro-American settlement. In such settings, paleoecology provides critical insights about past ecosystem responses to a range of climate conditions and disturbance regimes in the absence of significant human interference (Millspaugh et al. 2000). Pollen records from across the Greater Yellowstone Ecosystem, for example, show that whitebark pine (Pinus albicaulis) was once more abundant and widely distributed than at present (Iglesias et al. 2015). During the early Holocene, when summers were warmer and drier and fires were more frequent than at present, this species grew at all elevations; its present restriction to high elevations is partly a result of the Holocene expansion of lodgepole pine (Pinus contorta; Fig. 1, top). Assessing whitebark pine’s future based only on its present climate distribution underestimates its tolerance to warm summers and fire, and its intolerance of competitors. Today, the species is also under threat from the introduction of a non-native blister rust and warmer-than-previous winters. These novel conditions justify the need for careful monitoring of whitebark pine and the evaluation of any change against its response range to past climate and disturbance.
Preserving biodiversity in cultural landscapes
Alpine landscapes have experienced a long history of human activity, ranging from intensive agriculture in the lowlands to pastoralism at high elevations. Many of these landscapes are culturally and economically significant and some are “iconic” examples of traditional land use (Fig. 1, bottom). Highly valued cultural landscapes in Europe include the heathlands of Western Europe, Mediterranean dehesas of Spain, sweet chestnut and cork oak forests of southern Europe, and high diversity pastures and dry meadows of Central Europe and the Alps. Defining a climate-based range of acceptable variability for these landscapes makes little sense, because fire regimes have long been altered by humans, who changed forest vegetation through introductions, cultivation or setting aside of species (Conedera et al. 2017).
For example, although the forests and grasslands in Switzerland are predominantly cultural landscapes, humans have and are altering their natural biodiversity differently. Alpine grasslands, prized for species richness, are now threatened by land abandonment and forest encroachment (Colombaroli and Tinner 2013). In contrast, forest diversity has been steadily depleted as a result of the deliberate reduction of some trees (e.g. Tilia, Fraxinus and Abies; Tinner et al. 2013) and the addition and promotion of others (Castanea, Taxus, Juniperus, Fagus, Picea; Conedera et al. 2017). The challenge for future conservation efforts in this region is to balance the legacies of past climate and land use against the conservation of cultural landscapes and ecosystems and their services.
Incorporating ecosystem history into conservation strategies
Recognizing the degree of past landscape alteration can help determine the appropriateness and feasibility of management objectives that seek to restore naturalness or protect cultural heritage. Between nearly pristine conditions at one end and highly altered conditions at the other, landscapes in the intermediate part of the gradient support both natural and cultural components (Fig. 2). These intermediate landscapes pose conservation challenges – to support natural structure and diversity and at the same time maintain cultural values – but they also represent conservation opportunities (Lindenmayer and Hunter 2010). For example, disturbance-sensitive species often grow as isolated, relict forests in a matrix of human-altered vegetation, facing high risks of extinction in some cases (e.g. Abies nebrodensis in Sicily; Fig. 2). Although the climate may be suitable for forest expansion, restoration requires fire suppression, grazing protection and removal of competing species to a level that may not be cost-effective or feasible (e.g. Henne et al. 2015). Deliberate intervention, careful management, and knowledge of landscape history can help guide realistic conservation planning.
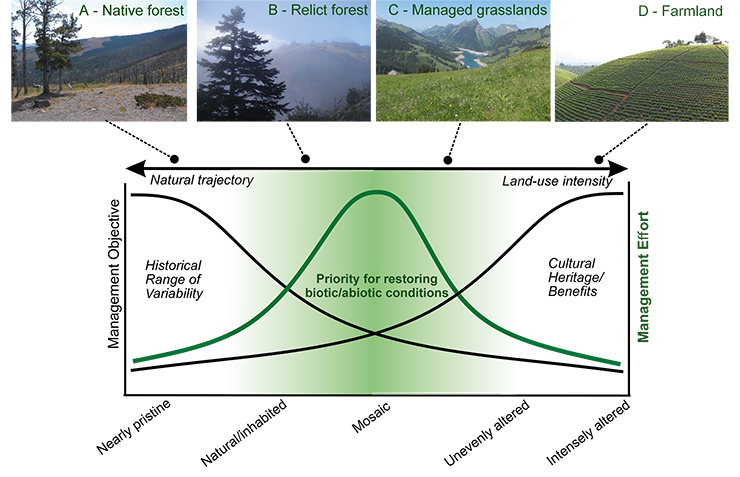 |
|
Figure 2: Top. Examples of landscapes with different levels of human alteration: (A) native Pinus contorta pine forest in Yellowstone National Park; (B) relict Abies nebrodensis forest in Sicily; (C) managed grasslands in the Alps; (D) tea crops in western Uganda (photos by DC). Bottom. Conceptual model showing management objectives across a gradient of landscape conditions, ranging from preserving historical range of variability in nearly pristine locations to promoting cultural benefits in intensely altered settings. The “landscape space” in the middle (green shading) highlights where pristine and humanized components co-exist and represent conservation opportunities for management objectives, for their potential to restore both historical range of variability and cultural values (based on Whitlock et al., in press). |
Paleoecological information can play a key role in evaluating current ecosystem states, supporting (or disproving) the effectiveness of present conservation measures to balance specific targets (Fig. 2), and assessing future management strategies and vulnerability to climate change (e.g. Tinner et al. 2013). Capturing environmental and ecological variability requires knowledge of past climate legacies and human impacts, although such information is unevenly distributed and absent in some regions (e.g. tropical rainforests; Colombaroli et al. 2016). Research to better understand past fire activity, as it relates to climate, people and biodiversity, is the focus of the PAGES Global Paleofire Working Group (pastglobalchanges.org/gpwg2).
affiliations
1Institute of Plant Sciences and Oeschger Center for Climate Change Research (OCCR), University of Bern, Switzerland
2UGent Limnology research unit, Ghent, Belgium
3Centre for Quaternary Research, Royal Holloway University London, UK
4Montana Institute on Ecosystems and Department of Earth Sciences, Montana State University, Bozeman, USA
5Swiss Federal Research Institute for Forest, Snow, and Landscape WSL, Cadenazzo, Switzerland
contact
Daniele Colombaroli: daniele.colombaroliips.unibe.ch
references
Colombaroli D et al. (2013) Divers Distrib 19: 157-170
Colombaroli D et al. (2016) J Trop Ecol 32: 213-225
Colombaroli D, Tinner W (2013) Holocene 23: 1625-1634
Conedera M. et al. (2017) For Ecol Manage 388: 100-112
Diffenbaugh NS, Field CB (2013) Science 341: 486-492
Gillson L et al. (2013) Trends Ecol Evol 28: 135-142
Henne PD et al. (2015) Front Ecol Environ 13: 356-362
Iglesias V et al. (2015) Plos One 10, e0124439
Lindenmayer D, Hunter M (2010) Conserv Biol 24: 1459-1468
Millspaugh SH et al. (2000) Geology 28: 211-214
Tinner W et al. (2013) Ecol Monog 83: 419-439
Williams JW, Jackson ST (2007) Front Ecol Environ 5: 475-482
Willis KJ, Birks HJB (2006) Science 314: 1261-1265
Whitlock C et al. (in press) Conserv Biol, doi:10.1111/cobi.12960
Peter Gell1 and Lindsey Gillson2
The decline in the biodiversity of the Earth is a critical global issue with potential impacts on essential ecosystem services. The rate of biodiversity loss has already far exceeded safe planetary boundaries (Rockström et al. 2009) and measures to arrest species extinction have been found wanting, with conservation reserves too small, too few and too subject to change. So, society has called for a significant increase in investment in biodiversity conservation, particularly given the additive impacts of accelerated climate change (Pecl et al. 2017). In concert, understanding of ecological function and process is needed if ecosystem services are to be managed sustainably.
Conservation biology has long been identified as the “crisis science” (Soulé 1985). However, preventing species extinction is not enough to maintain ecological function, socio-ecological resilience and evolutionary potential (Sgrò et al. 2011). The links with society and the sustainable use of landscapes are increasingly part of the broader conservation remit (Wu 2011). The magnitude of the issues encountered greatly exceeds the foundational ecological knowledge available, and counting species and population declines is not enough. Historically, conservation biologists have grappled with the design of protected area networks, and preventing deforestation and desertification. Today, the portfolio of challenges has expanded to include ecological isolation, fragmentation and degradation of terrestrial and aquatic systems, and accommodating species whose habitats are at risk from changing climate patterns, as well as the sustainable use of natural resources. Furthermore, maintaining and restoring appropriate disturbance regimes has become increasingly important as land use has pushed disturbance regimes beyond their historical limits (North and Keeton 2008).
 |
|
Figure 1: The Gamo Highlands of Ethiopia – a landscape shaped by multiple interacting drivers (Photo: Rob Marchant). |
In fact, the rate of biodiversity loss tempts conservation biologists to focus predominantly on the recent, and even just the now. Here, however, they run the risk of misunderstanding the landscape elements that influenced system function, the factors that drove the declines and the historical role of people as ecological architects (Fig. 1). Further, they may overlook the existence of cultural ecosystems and the past, and future, role of people in them (Wu 2011). Yet even the value of these lessons may be challenged, as “novel ecosystems” emerge due to anthropogenic climate change, land use and alien species (Williams and Jackson 2007).
We maintain that the study of long-term ecosystem change can provide critical insights into the resilience of ecosystems, the restoration of disturbance regimes and the prediction of future responses to changing climate and disturbance – even when conditions are novel. This collection of papers is an illustration of this view.
The role of disturbance in ecosystems is widely recognized but rarely is the legacy of historical disturbance, particularly by humans, acknowledged in conservation planning. The neo-tropical Gran Sabana is a mosaic of vegetation that reflects the influence of climate change but also human burning as an agency of vegetation dynamics (Rull et al., p.82). The management of the vegetation mosaics here, as well as in the North American pine communities, requires an appreciation of the influence of cultural practices, and the impact of their removal (Colombaroli et al., p.78). The east African landscapes too, that have supported diverse megafaunas in close association with long human habitation (Marchant et al., p.80), attest to long interactions between people and biodiversity, but with this balance challenged in recent decades by the intensification of development.
The legacy of human occupation is often an increase in regional biodiversity, and cultural landscapes are themselves now considered valid conservation targets. The contemporary richness of some European communities evolved in association with human disturbance (Ekblom and Gillson, p.88). Similarly, the legacy of horticulture is the augmentation of botanical diversity in long-reforested Central and South American regions (Whitney and Cárdenas, p.84).
These vignettes of human-ecosystem interactions attest to the importance of long-term evidence to inform appropriate landscape management. Rarely has this occurred, however UN programs such as the Ramsar Convention are recognizing long-term variability to better ascribe natural ecological character and limits of acceptable change (Gell, p.86). Understanding ecological responses to external, including human, stressors is critical if management efforts are to be well targeted (Seddon, p.94). The scaling up of paleoecological records represents a means by which long-term data (Harrison, p.96) could be brought to biodiversity management decision making.
This issue of Past Global Changes Magazine focuses on the perspectives that can be brought to the crisis science from paleoecological evidence. It draws on lessons from the Americas, Africa and Europe, while also providing new insights into the long-term geological (Fritz and Baker, p.90) and climatic (Ledru et al., p.92) forces of biodiversity evolution, critical transitions in ecosystem change (Seddon, p.94) and utility of ecosystem change evidence for global conservation programs to enable them to better address the challenges imposed by intensive development. Lastly, it advocates for big data as a means of scaling up our knowledge to contribute best to informing programs that address biodiversity management (Harrison, p.96).
affiliations
1Water Research Network, Federation University, Ballarat, Australia
2Department of Biological Sciences, University of Cape Town, Rondebosch, South Africa
contact
Lindsey Gillson: lindsey.gillsonuct.ac.za
references
North MP, Keeton WS (2008) In: Lafortezza R et al. (Eds) Springer, 341-372
Pecl GT et al. (2017) Science 355: eaai9214
Rockström J et al. (2009) Nature 461: 472-475
Sgrò CM et al. (2011) Evol Appl 4: 326-337
Soulé ME (1985) Bioscience 35: 727-734
PAGES 2k Network Coordinators*
Introduction
The past 2000 years (the “2k” interval) provides critical context for our understanding of recent anthropogenic forcing of the climate, as well as baseline information about Earth’s natural climate variability. It also provides opportunities to improve the interpretation of paleoclimate proxy observations, and to perform out-of-sample evaluation of the climate and earth system models that are used to generate projections of future climate change.
Achievements of the PAGES 2k Network
 |
|
Figure 1: 30-year-mean temperatures for the seven PAGES 2k Network regions, standardized to have the same mean (0) and standard deviation (1) over the period of overlap among records (1190–1970 CE). North America includes a shorter tree-ring based and a longer pollen-based reconstruction. Adapted from PAGES2k Consortium (2013). |
In 2008 PAGES initiated the 2k Network, to coordinate and integrate regional efforts to assemble existing proxy data and generate climate reconstructions. Nine regional groups were established during the course of the initiative, spanning eight continents and the global ocean. Phase 1 (2008-2013) focused on generating regional temperature reconstructions. During Phase 2 (2014-2016), as a natural step forward, a number of trans-regional groups emerged from amongst the community, focusing on topical challenges such as methods development, data-model comparison, database construction and large-scale climate.
Along with the many products and publications of the regional groups, the initiative spawned a number of successful, large network-wide projects. The first key product was the coordinated publication of temperature reconstructions from seven continents, which informed the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (PAGES 2k Consortium, 2013). For the first time, continental-scale temperature histories spanning the 2k interval were systematically compared. The results showed that there were no globally-consistent temperature fluctuations consistent with a worldwide “Medieval Warm Period” or “Little Ice Age” (Figure 1). However, there was a near-global long-term cooling trend during the Common Era, which culminated in a cold interval from 1580 to 1880 CE. In contrast, the period from 1971 to 2000 CE was the warmest during the last 1400 years. Through this publication, the 2k Network received considerable attention outside the field of paleoclimate and amongst the general public and it remarks a milestone in making 2k paleoclimate science accessible and understandable to a wider audience. It currently ranks in the 99th percentiles of Earth and Planetary Sciences articles of the same age and document type in terms of citations, Mendeley readers and tweets, and it received 27 mass media mentions (www.scopus.com).
 |
|
Figure 2: Global SSTs over the last 2000 years: A cooling over the past two millennia was reversed only in the most recent two centuries. Fifty-seven previously published and publicly available marine sea surface temperature reconstructions were combined and compiled into 200-year brackets, represented by the boxes. The thin horizontal lines dividing each box are the median of the values in that box. The thick blue line is the median of these values weighted for differences in the region of the global ocean in which they were found. Modified from McGregor et al. (2015). |
In collaboration with the Paleoclimate Modelling Intercomparison Project (PMIP), these regional reconstructions were compared with transient simulations of the last millennium (850 to 1850 CE). The resulting publication identified a consistency in the general tendencies, but temperature changes in different regions correlated more closely with each other within the simulations than within the reconstructions (PAGES2k-PMIP3 group, 2015). Subsequent efforts by the Ocean2k Working Group found that a robust long-term cooling trend also occurred over the global ocean during the pre-industrial Common Era (Figure 2; McGregor et al., 2015). Comparison with climate model simulations suggested that this trend was driven by clusters of volcanic eruptions.
 |
|
Figure 3: Onset of industrial-era warming in regional temperature reconstructions. Regional reconstructions since 1500 CE (coloured lines) with 15-yr (thin black lines) and 50-yr (thick black lines) Gaussian smoothing, shown alongside the median time of onset for sustained, significant industrial-era warming assessed across 15–50-yr filter widths (vertical red bars). Grey 1 °C scale bar denotes the y-axis scale of each regional temperature reconstruction. Modified from Abram et al. (2016). |
Combination of the continental reconstructions with temperature histories from ocean basins over the last 500 years yielded the curious finding of an early onset of industrial-era warming, which could already be detected in the mid-19th century over the tropical oceans and Northern Hemisphere continents (Figure 3; Abram et al., 2016). This trans-regional effort highlighted that the effect of greenhouse-gas forcing on temperatures started earlier than is suggested by instrumental data alone. Additionally, a spatial reconstruction of precipitation across the Northern Hemisphere for the past 1200 years, in comparison with climate model simulations, suggests that models do not yet accurately simulate long-term hydroclimate variability (Ljungqvist et al., 2016). Finally, a community-wide effort of Phase 2 has been to systematically organize temperature-sensitive proxy data and metadata covering the 2k interval into a common database product, to facilitate future assessments of temperature variability during this period.
 |
|
Figure 4: Quality screened proxy records assembled in the PAGES2k temperature database at the end of Phase 2 (PAGES 2k Consortium, in press). Number of records per archive is indicated in the legend. Background image from http://visibleearth.nasa.gov. |
Common to all these community products has been a tremendous collaborative effort involving hundreds of scientists from all regions of the globe, aiming to improve our understanding of mechanisms of climate variation on interannual to bicentennial time scales by contributing expert knowledge, data and metadata. As an example of the impressive progress in data collection and synthesis, Figure 4, shows the current availability of quality-screened temperature-sensitive proxy data (PAGES 2k Consortium, in press).
Our vision for Phase 3
The goals of Phase 3 (2017-2019), which was launched in May 2017 at the PAGES Open Science Meeting in Zaragoza, Spain, are to:
1. Further understand the mechanisms driving regional climate variability and change on interannual to centennial time scales (Theme: “Climate Variability, Modes and Mechanisms”);
2. Reduce uncertainties in the interpretation of observations imprinted in paleoclimatic archives by environmental sensors (Theme: “Methods and Uncertainties”);
3. Identify and analyse the extent of agreement between reconstructions and climate model simulations (Theme: “Proxy and Model Understanding”).
Research is organized as a linked network of well-defined projects and targeted manuscripts, identified and led by 2k members. This bottom-up concept of trans-regional projects initiated and conducted by community members has successfully emerged during Phase 2 and is expected to further stimulate collaboration within the 2k Network. The 2k projects focus on specific scientific questions aligned with Phase 3 goals, rather than being defined along regional boundaries. Along with the products of the individual projects, one or more community wide projects are envisaged for Phase 3.
As mentioned above, an enduring element from earlier phases of PAGES 2k will be a culture of collegiality, transparency, and reciprocity. Phase 3 seeks to stimulate community based projects and to facilitate collaboration of researchers from different regions and career stages, drawing on the breadth and depth of the global PAGES 2k community. A key vision of PAGES 2k is to support end-to-end workflow transparency, open data and knowledge access, which are key conditions for the inclusion of future PAGES 2k projects. The initiative seeks to further develop collaborations with other research communities and engage with stakeholders at the project and network level through interaction with related institutions or initiatives such as Future Earth, WCRP, IPCC and EarthCube.
Call for participation
There are many ways to participate in, and to be part of, the PAGES 2k community. You may contribute to the ongoing database and knowledge-base efforts with your data and expertise. You may initiate a new 2k project, or participate in an emerging one, by contributing towards project coordination, data-analysis, interpretation or writing. In the spirit of Phases 1 and 2, PAGES 2k projects are expected to be inclusive and open to any researcher who wishes to contribute. Members from related communities including CLIVAR scientists are warmly welcomed to be part of or even initiate 2k projects.
If you would like to participate in Phase 3 of the PAGES 2k Network or simply to receive updates, please visit http://www.pastglobalchanges.org/ini/wg/2k-network/intro to join our mailing list or contact a coordinating committee member. A call for new PAGES 2k projects will soon be issued via the mailing list.
affiliations
PAGES IPO, Falkenplatz 16, Bern, Switzerland
references
*PAGES2k Coordinators
Nerilie Abram, The Australian National University, Canberra, Australia, nerilie.abramanu.edu.au
Oliver Bothe, Helmholtz-Zentrum Geesthacht, Geesthacht, Germany, ol.bothegmail.com
Hans Linderholm, University of Gothenburg, Göteborg, Sweden, hanslgvc.gu.se
Belen Martrat, Spanish Council for Scientific Research, Barcelona, Spain, & University of Cambridge, Cambridge, UK, belen.martratidaea.csic.es
Helen McGregor, University of Wollongong, Wollongong, Australia, mcgregoruow.edu.au
Raphael Neukom, University of Bern, Switzerland, neukomgiub.unibe.ch
Steven Phipps, University of Tasmania, Hobart, Australia, Steven.Phippsutas.edu.au
Scott St. George, University of Minnesota, Minneapolis, USA, stgeorgeumn.edu
Lucien von Gunten, PAGES International Project Office, Bern, Switzerland, lucien.vonguntenpages.unibe.ch