PAGES Magazine articles
Frank M. Chambers![]()
For degraded mires and moorlands in Europe, paleoecological data from peats reveal how and when the present landscape developed. These data widen the vision of future landscapes and legitimate a greater range of targets for nature conservation agencies.
Context
The timescales and imperatives of scientists reconstructing past environments (paleoecologists), and those involved in modern ecology, have been very different (Birks and Birks 1980). Data produced by the former may be unread by, or unintelligible to, the latter, owing to the specialist journals in which they are published and their manner of presentation. Attempts to "bridge the gap" (Davies and Bunting 2010) between paleoecologists and ecologists are infrequent. Examples can be found in environments for which detailed paleoecological data are available to chronicle the past, showing how the present landscape developed (Gillson 2015). There is great potential to include these studies in restoration ecology.
Mires and moorlands
In Europe, while ecosystems responded to climate changes during the Holocene, many have also been affected by human activity over millennia. This is not always fully recognized by those responsible for their conservation. For example, in British mire and moorland environments, the most obvious human-induced effect of loss of Sphagnum (bog moss) species over the last century is well-known from naturalists' records, and attributed to pollution from sulfur dioxide. Yet, the long-term effects of centuries of grazing animals, at different stocking rates, with different species, is often ignored or unappreciated.
 |
|
Figure 1: Stainless-steel peat-cutter, showing safety-sheath (left) and field use (center); its sharp edge (right) ensures a clean cut, and minimal environmental damage. |
Mires and moorlands have the advantage that their underlying peats, which are incompletely decayed accumulations of plant remains, contain a record of their vegetation development over time. Analysis of these remains—pollen, spores, plant macrofossils—and of other properties of the peat, allows for reconstruction not only of their vegetation through time, but also of past climate change (Barber et al. 1994). Careful framing of multiple hypotheses permits inference of the relative effects of human activity on these landscapes.
Many nature conservationists may be unaware of the previous natural or semi-natural states of environments, beyond the past few decades. Indeed, 1949, when the national conservation agency was first established in the UK, has been used as a definitive baseline for comparative purposes of percentage habitat loss and degradation. Fortunately, enlightened staff in some of the national agencies have recognized the potential for paleoecologists to contribute much longer-term data to inform contemporary and future conservation. Nevertheless, assimilation and use of these data by conservation practitioners is not easy, requiring a more accessible form of presentation of paleovegetation data, particularly replacements for intricate pollen diagrams.
First overtures
An early attempt to combine the use of paleoecology with contemporary research into moorland restoration commenced on Exmoor, southwest England, in the late 1990s. The contemporary work, reported later by Marrs et al. (2004), but without reference to the paleo studies, had set out experimental plots investigating the use (or not) of herbicides, summer grazing (or not), and their use in combination to control and reduce the dominance of purple moor grass (Molinia caerulea). This dominance was regarded as undesirable in this designated Environmentally Sensitive Area (ESA), both on aesthetic grounds—the plant produced monotonous landscapes in a much-visited national park—and because it offered unpalatable grazing for stock, principally sheep. It was assumed, based on contemporary observations and oral accounts, that the purple moor grass had recently supplanted heather (Calluna vulgaris)—a plant regarded as aesthetically pleasing.
Palaeoecological data confirmed the relatively recent spread and expansion of purple moor grass, but unexpectedly revealed that the landscape had alternated between grass moor (with purple moor grass probably only a minor component) and heather moorland over the past millennium (Chambers et al. 1999). These results legitimized present attempts to control the hitherto unprecedented spread and dominance of purple moor grass but offered different visions of what the landscape could look like. Combining the two separate lines of research (experimental plots and paleoecology) suggests that generalist-feeding ponies or cattle, rather than sheep (who are finicky feeders), might help to reduce the dominance of purple moor grass.
The hitherto unprecedented stocking levels of sheep could, in part, explain the recent decline of heather and its replacement by Molinia. Speculatively, the combination of previous land management and grazing practices, combined with climate changes through the Medieval Climate Anomaly and the Little Ice Age, might explain the previous alternations between grass moor and heather moorland. Thus, the vision of the future landscape, upon restoration from its current monotonous state, does not necessarily have to be the replacement of one dominant (Molinia) by another (Calluna): a more diverse mosaic of grass–heather moor could be envisaged.
 |
|
Figure 2: Paleoecological data (selected taxa) from Brecon Beacons, South Wales, showing rise and fall of Sphagnum (green arrows), and recent rise of purple moor grass (Molinia) after the start of the Industrial Revolution (14 cm depth). |
Degraded mires in South Wales
In South Wales, it was acknowledged that many upland mires and moorlands are depauperate, lacking some typical mire plants, particularly some of the bog moss species. This was variously attributed to overgrazing, too-frequent burning, airborne pollution, peat erosion, and climate change, among other causes. In many localities, Molinia was dominant.
The amount of carbonaceous pollution that fell on the uplands since the start of the Industrial Revolution had been roughly calculated (Chambers et al. 1979), and there had been the assumption that sulfurous atmospheric pollution was largely to blame for loss of Sphagnum species, as in the Pennines (Lee et al. 1987). However, any associated nutrient loading, which might give competitive advantage to Molinia, was hardly considered, and is not easy to assess. Other sources of eutrophication might be long-distance nitrogen deposition associated with agricultural practices, and the fertilizer effect of droppings from sheep at high stocking levels being fed winter feed supplement, which could result in nutrient enrichment of uplands.
On the Brecon Beacons, the current landscape is mainly treeless moorland and blanket bog. Some of the latter is eroding, with upstanding haggs (eroding blocks) of peat. The vegetation lacks diversity, with areas dominated variously by cotton sedge or purple moor grass. Palaeoecological research, commissioned by the Countryside Council for Wales (CCW; now Natural Resources, Wales), was undertaken to understand how the landscape had developed—not over the last few decades, but over the last 500 years. Radiocarbon dating showed that the peat had accumulated over several thousands of years, and so, with permission, the story was extended by taking peat monoliths economically using special field equipment (Fig. 1). The analysis of deep peat sections proved particularly useful, as it put the current vegetation state into a much longer time perspective (Chambers et al. 2013). The data showed that the current vegetation had no longevity, but rather had developed since the Industrial Revolution, with the rise of purple moor grass a recent feature after decline of centennial (but not millennial) dominance of Sphagnum (Fig. 2), suggesting a range of possible restoration targets.
Recent and future collaboration
In the decade since an earlier review in this magazine (Chambers and Daniel 2011), there has been limited application of palaeoecological data for mire and moorland conservation in the UK. A notable exception has been the Yorkshire Wildlife Trust, through the Yorkshire Peat Partnership, who sponsored research by McCarroll et al. (2016; 2017) on selected mires to understand better how to effect mire restoration. In Wales, too, earlier work co-published with CCW staff (Chambers et al. 2007) helped to inform conservation practice. These are exceptions rather than commonplace, and though a timely and potentially useful review of the utility of palaeoecological techniques for moorland management is available (Table 6 in Chambers et al. 2017), it has been scarcely sighted by conservation practitioners, and rarely cited. One possible reason why much mire and moorland restoration and management does not involve preceding paleoecological research is that, even with medium-term (5-yr) funding, conservation agency targets seem immediate or short-term compared with datasets covering hundreds or thousands of years. If longer-term goals were set, paleoecological research could be aligned and the long-term data generated could be better assimilated to inform the trajectory.
affiliation
Centre for Environmental Change and Quaternary Research, School of Natural and Social Sciences, University of Gloucestershire, UK
contact
Frank M. Chambers: fchambers glos.ac.uk
glos.ac.uk
references
Barber KE et al. (1994) Holocene 4: 198-205
Birks HJB, Birks HH (1980) Quaternary Palaeoecology. Arnold, 289 pp
Chambers FM, Daniell JRG (2011) PAGES News 19: 45-47
Chambers FM et al. (1979) Nature 282: 829-831
Chambers FM et al. (1999) J Appl Ecol 36: 719-733
Chambers FM et al. (2007) Biodivers Conserv 16: 2821-2846
Chambers FM et al. (2013) Biodivers Conserv 22: 719-736
Chambers FM et al. (2017) AIMS Environ Sci 4: 54-82
Davies AL, Bunting MJ (2010) Open Ecol J 3: 54-67
Marrs RH et al. (2004) J Appl Ecol 41: 398-411
K. Anggi Hapsari![]() 1, T.C. Jennerjahn
1, T.C. Jennerjahn![]() 2,3 and H. Behling
2,3 and H. Behling![]() 1
1
There are several potential practical applications of paleoecological information that can provide guidance for improving peatland restoration in Indonesia, and highlight the values, roles, and applicability of paleoecology in ecological restoration.
The importance of peatlands in Indonesia
Indonesian peatlands provide habitat for a large diversity of organisms, including endangered flora and fauna. They also store ∼56 Gt carbon, equivalent to five years of current global carbon emissions, mainly in their soggy, spongy soil layers made of partially decomposed dead plant material. In the past few decades, rapid peatland degradation and drainage (Fig. 1) has not only posed a threat to the native orangutans and other species living there, who are rapidly losing their habitat, but has also facilitated peat fires, leading to haze and related health crises, and costing Indonesia billions of dollars in economic loss (World Bank 2019). Peat fires also released enormous amounts of carbon dioxide, fueling climate warming, and placing Indonesia amongst the world's top carbon emitters. It is not surprising there are growing calls for restoration.
 |
|
Figure 1: Images of (A) pristine, (B) degraded, and (C) burnt peatlands in Indonesia (photo credits: A. Hapsari and I. Fikri). |
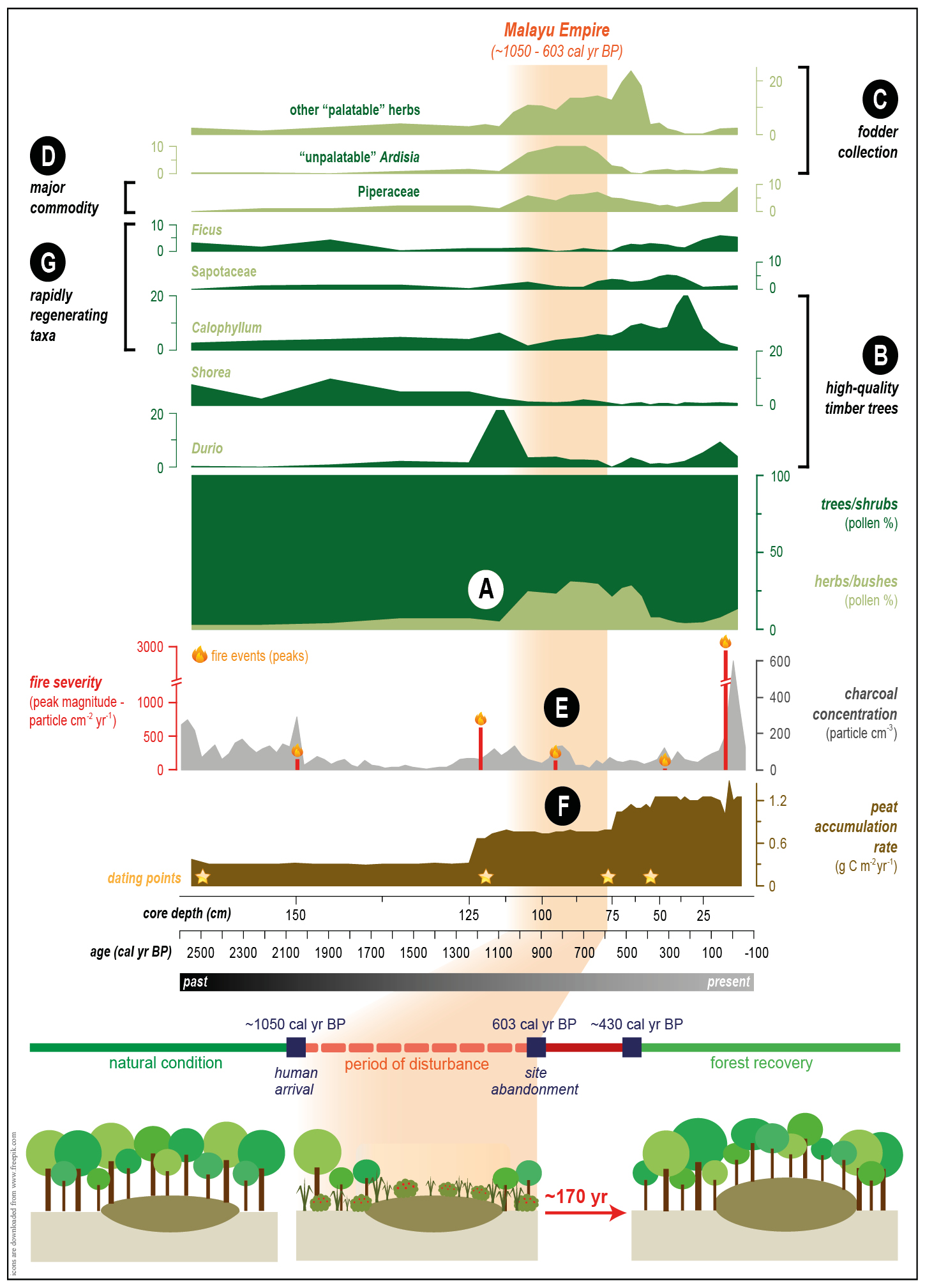
"Ecological restoration can be improved using paleoecology": a statement easily found in many paleo-related publications. However, there is a dearth of examples on how paleo-information can be practically applied in restoration beyond defining pre-human-intervention baselines. Meanwhile, attempting to restore the ecosystems to their pre-human conditions is increasingly being considered to be unattainable or even inappropriate (Clewell and Aronson 2013; Baker and Eckerberg 2016; Rull, this issue). Here, we use a paleorecord from the Sungai Buluh peatland in Indonesia as our main example, where the impact of the Malayu Empire's activities and how the peatland responded to them, is documented (Fig. 2; Hapsari et al. 2018; 2021).
Goal adjustment
When defining what it is we are trying to restore, we can use ecological attributes of a successfully restored or undegraded ecosystem (e.g. species composition, resilience, functions) as goals (Clewell and Aronson 2013). We then need to ensure the goals are realistic and can be met within regulatory constraints and available resources.
Most, if not all, peatland restoration projects in Indonesia aim to restore both hydrology and peat swamp forests to an intact state, i.e. prior to disturbance, and refurbish their functions and values (Puspitaloka et al. 2020). These projects are typically planned for 60 years, but many are shorter (3-30 years) due to limited resources.
The paleorecord from Sungai Buluh taught us that it took 170 years for the peatland to return to its pre-disturbance condition after "light" human use (i.e. no clearing/drainage). Thus, the goals of present-day peatland restoration are quite unrealistic, as we now face much harder ecological (drained/cleared/burnt) and socioeconomic (resistance/sabotage) challenges, a much shorter timeframe, and often limited resources. It is necessary for the projects to either opt for easier goals (e.g. zero-illegal logging, blockage of all canals) or to extend the project durations, especially if the fulfillment/unfulfillment of the set goals will decide the project's fate, e.g. further funding, permit extension, or penalty.
 |
|
Figure 2: The Sungai Buluh peatland underwent major changes throughout the Malayu Empire occupation (orange shade; modified from Hapsari et al. 2018; 2021): (A) Peat swamp forest cover declined soon after the Malayu Empire arrival (∼1050 cal yr BP); (B) A lower proportion of high-quality timber trees and historically recorded natural resource use for construction by the Malayu Empire suggests selective timber collection; (C) Vigorous growth of unpalatable forage that is usually driven by the lack of competition from other palatable herbs suggests fodder collection; (D) An increase in Piperaceae while the Malayu Empire became the largest pepper center in Sumatra implies non-timber forest product (NTPF) collection; (E) Relatively low charcoal concentration and peak magnitude suggests inextensive fire use; (F) Continuously accumulating peat substrate that needs a consistently high water-table suggests an absence of land drainage; (G) PSF regeneration after ME abandonment started with the increase of rapidly regenerating taxa. |
Refine the designs
The Sungai Buluh record suggests that a peatland can fully recover if it isn't drained and still has some of the old forest remaining. Restoration projects for peatlands with these conditions can choose the low-cost prescribed natural regeneration (passive restoration) approach. The resources available can be concentrated to remove/reduce the disturbances, e.g. fire prevention, law enforcement, ecoliteracy and bioeconomy development, or alternative livelihood creation.
For the drained and deforested peatlands, assisted natural regeneration or partial/complete reconstruction is needed, which includes replanting and/or rewetting. However, although replantation in peatlands creates an alternative livelihood for the locals, it is very expensive and often fails because unsuitable taxa are used, e.g. dryland species are planted in deeply flooded conditions (canal/burnt scar). A paleorecord from Borneo shows that peatlands sometimes developed in Pandanus-dominated freshwater lakes (Hope et al. 2005). Replantation can then mimic natural succession by planting Pandanus in the deeply flooded areas (lake-like conditions) and introduce other desirable taxa once those areas are shallow enough (Giesen and Sari 2018).
For restoration to be successful, the target ecosystem needs to eventually manage to self-organize, an attribute that is mainly driven by the planted taxa's regenerating ability. The recovery of the Sungai Buluh peat swamp forest started with recruitment of taxa with high seed productivities, viabilities, germination rates, and seedling survival rates. Peatland restoration plans thus need to include those and/or other taxa with similar regenerative power and, if necessary, to introduce specific pollinators or seed dispersers (e.g. Trigona bees for Shorea).
Paleorecords also suggest that dense forest cover is the secret of peatlands' fire-resistance (Hapsari et al. 2021). To improve target peatlands' resilience to fire, forest cover needs to be grown quickly. While the current strategy chooses to plant in single-species groups, introducing a few individuals from many species and letting the "selection effect" do the work is a better option than planting a large number of poorly adapted species and having none survive.
Stream of income
In restoration, socioeconomic achievement is important, but it won't be sustainable long-term without ecological success and durability. Subsequent management of restored peatland needs to ensure that the socioeconomic activities won't jeopardize peatland resilience. The Sungai Buluh record suggests that collection of non-timber forest products and fodder, and selective/restrictive wood extraction without draining the land or burning the trees, are sustainable. A paleorecord from Borneo (Hope et al. 2005) also shows that human impact was restricted to the riversides because of access difficulties. Restoration can mimic the low level of past human disturbance by concentrating economically valuable taxa alongside natural waterways to ease people's access to their income source, thus limiting the temptation to reopen blocked canals, which cause major challenges in peatland rewetting.
Time is of the essence
Restoration, unfortunately, isn't a quick fix, but a long-term effort that requires patience and dedication (Clewell and Aronson 2013). Policy in Indonesia constrains the durations of peatland restoration projects within 60 (extendable to 100) years. Assuming that target peatlands will also require about 170 years to recover, like the Sungai Buluh, the 60-year framework is far too short. In fact, the target peatlands are often drained, thus requiring physical environment modification (rewetting), which will likely prolong the recovery time.
A major driver for restoration success is the time that has elapsed since the restoration started (Crouzeilles et al. 2017). Thus, the current policy needs to allow for some flexibility in granting permissible periods according to the projects' goals, e.g. longer duration for projects targeting a full biodiversity recovery than for projects aiming at zero-carbon emission (Hapsari et al. 2018). If the current policy remains in place, it needs to, at least, consider the substantial time lag between the completion of project tasks and the attainment of restoration targets. The 60(–100)-year permissible project length can serve as an "intervention period"; however, the restoration site should remain protected and monitored for another 60(–100) years to accommodate the development of peatlands' self-organization and self-sustainability.
A hope… and a warning!
Many people wonder whether ecological restoration is really achievable, or if it is just empty promises and a massive waste of investment (Conniff 2015; Almassi 2017). As the success of these efforts remains to be seen, the long wait may give rise to growing scepticism (Giesen and Sari 2018; Miller et al. 2021). The documented recovery of the Sungai Buluh peatland from past human disturbance certainly gives hope for tropical peatland restoration efforts as it shows that recovery is a slow process and that decades of waiting do not necessarily mean failure. However, it should also be taken as a cautionary example that it will take a whole lot longer to depave a parking lot and build back a paradise than the other way around—as in Joni Mitchell's song.*
*Big Yellow Taxi: jonimitchell.com/music/song.cfm?id=13
affiliations
1Department of Palynology and Climate Dynamics, University of Göttingen, Germany
2Department of Biogeochemistry and Geology, Leibniz Centre for Tropical Marine Research (ZMT), Bremen, Germany
3Faculty of Geoscience, University of Bremen, Germany
contact
K. Anggi Hapsari: kartika.hapsaribiologie.uni-goettingen.de
references
Almassi B (2017) Ethics Environ 22: 19-40
Baker S, Eckerberg K (2016) Restor Ecol 24: 284-290
Conniff R (2015) The false promise of ecological restoration projects. TakePart Magazine
Crouzeilles R et al. (2017) Sci Adv 3: e1701345
Hapsari KA et al. (2018) J Ecol 106: 1-18
Hapsari KA et al. (2021) Rev Palaeobot and Palynol 293: 104482
Hope G et al. (2005) Quat Res 64: 407-417
Miller MA et al. (2021) Sustain Dev 1-15
Katarzyna Marcisz![]() 1, S. Czerwiński
1, S. Czerwiński![]() 1,2, M. Lamentowicz
1,2, M. Lamentowicz![]() 1, D. Łuców
1, D. Łuców![]() 1,3 and M. Słowiński
1,3 and M. Słowiński![]() 3
3
As one of the worlds' most important carbon stocks, peatlands must be protected and restored. Paleoecology can be regarded as an important tool in peatland restoration and management; as decision making is a complex and intricate task, it should consider the long-term perspective of ecosystem development.
Peatlands' role in climate change mitigation
Peatlands cover 3% of the world's land area and store one-third of global terrestrial carbon, which makes them one of the most important carbon stocks (Rydin and Jeglum 2013). Saturated conditions in peatlands protect stored carbon. Healthy peatlands are also key players at the ecosystem level because they function like sponges and accumulate water in the landscape (Rydin and Jeglum 2013). Hence, they positively affect adjacent ecosystems, such as forests or grasslands.
Most peatlands worldwide have experienced significant human pressure in the past, mainly through peat extraction (Kołaczek et al. 2018) or various forms of drainage (Talbot et al. 2010), which led to the lowering of the water table. The drying of European peatlands has intensified over the last 300 years (Swindles et al. 2019), and the proportion of degraded peatlands in Europe is high (Tanneberger et al. 2021). Conservation is, therefore, crucial to prevent peatlands from turning from carbon sinks to carbon sources and to help store more water in the currently drying and warming world. Moreover, the maintenance of resilient peatlands will reduce ongoing biodiversity loss (Rydin and Jeglum 2013).
Assessing reference conditions using peatland paleoecology
Looking into the past by reconstructing long-term environmental changes is crucial to improve reference conditions for nature protection (Valsecchi et al. 2010), environmental and forest management (Hennebelle et al. 2018; Słowiński et al. 2019), and peatland restoration (Łuców et al. 2022). Reconstructed vegetation changes, hydrological fluctuations, and disturbance records (e.g. fires, human activity) can help determine potential vegetation composition, assess the extent of human impact, and single out factors that led to main vegetation transitions.
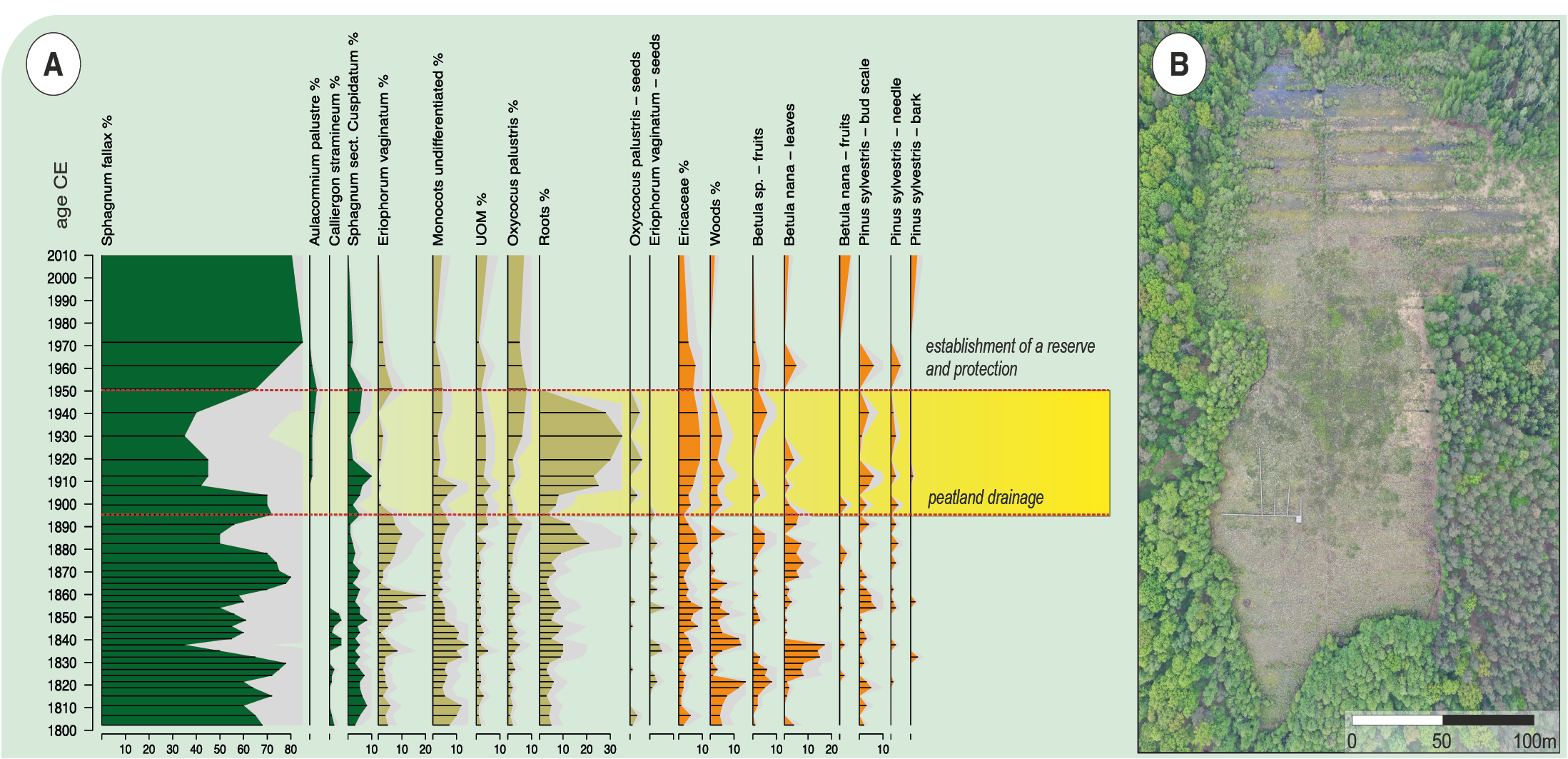
Human impacts have long-lasting consequences that are visible in palaeoecological records. For example, the effect of the establishment of drainage ditches on the Linje poor-fen (northern Poland) had immediate consequences on the local vegetation (Marcisz et al. 2015), and drainage ditches and remains of exploitation ponds are still visible on the site more than a century after drainage ceased (Fig. 1).
 |
|
Figure 1: Linje poor-fen located in northern Poland: (A) paleoecological view of the effect of melioration and peat extraction at the end of the nineteenth century and (B) drone photo of the site taken in 2021 showing still visible areas of the drainage ditches. |
Peatland histories are complex and very dynamic, and it is sometimes difficult to identify reference conditions in the past that can be set as a target for nature restoration. A paleoecological study of the Kazanie fen (western Poland), an alkaline fen that is now under restoration, shows a diverse history of a wetland that was affected by humans over the last millennium (Czerwiński et al. 2021). Deforestation in the surrounding catchment affected its hydrology and trophic state, leading to accelerated terrestrialization of the wetland and the formation of an alkaline fen. Overexploitation of the surrounding forest, related mostly to deforestation, led to the loss of water in the catchment, which was followed by climate warming, drying and, in the end, acidification of the site. In reality, it is the anthropogenic novel ecosystem of the alkaline fen that is the target of ecological restoration (Czerwiński et al. 2021). A similar case is the Pawski Ług bog (western Poland), which is now protected as a Nature Reserve. This peatland had been functioning as an alkaline fen until ca. 700 years ago, when it switched to an acidic bog. The change was an effect of deforestation of the surrounding forests and an introduction of a feudal economy by the Knights Hospitaller (a medieval and early modern Catholic military Order of Knights of the Hospital of Saint John of Jerusalem, also known as the Joannites) who settled in the area (Lamentowicz et al. 2020). Therefore, the bog that is currently protected is, in fact, a novel anthroecosystem that is far from its pristine state.
Peatland restoration efforts
In choosing proper restoration techniques, it is important to recognize the setting of the peatland, its catchment, and the factors influencing its development. Knowledge about peatland history can help implement a suitable restoration plan (Fig. 2). Yet, searching for an ecological baseline for peatland restoration may not be easy, and the efforts put into the restoration process may not result in the restoration of peatlands back to their former, pre-disturbed conditions (Łuców et al. 2022). Peatlands worldwide were influenced by various disturbance factors, and often the main one—human activity—was not present to the current extent and magnitude in the past. Therefore, it may be very hard to achieve satisfactory results in terms of biodiversity and vegetation composition.
 |
|
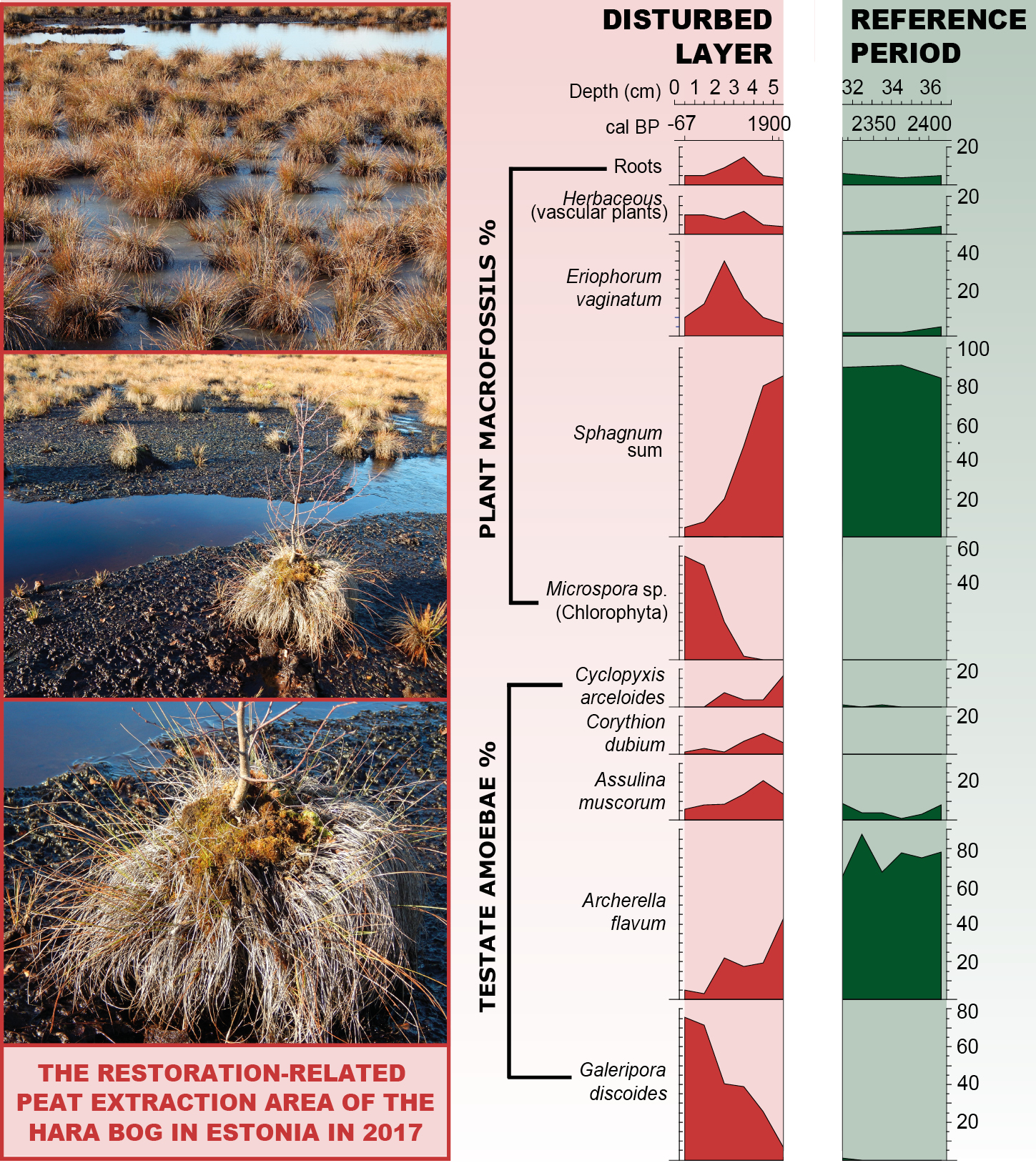
Figure 2: The restoration-related peat extraction area of the Hara bog located in northern Estonia. (Left) Pictures presenting the current structure of vegetation with hummocks of Eriophorum vaginatum and Sphagnum mosses, indicating fluctuations in the water table. (Right) Paleoecological record showing ecological contrast between restored state (current state) and pristine conditions (dated to ca. 2000 years before present) that can be regarded as a reference period for the restoration. Testate amoeba and plant macrofossil data suggest that, even though the Hara bog is far from stable and saturated conditions, it has the potential to be restored. However, the restoration of peat vegetation on bogs is a complex, difficult, and time-consuming process (figure modified from Łuców et al. 2022). |
The most common technique used for peatland restoration is rewetting drained sites using various types of dams constructed on ditches. Rewetting helps reinstate high moisture levels on the peatland and create bog-like conditions (Hancock et al. 2018), which protects carbon stored in the deeper peat and can reduce the number of peat fires (Sirin et al. 2021). However, even though novel ecosystems created through such interventions may not resemble previous peatland vegetation (Kreyling et al. 2021), e.g. be dominated by more common Sphagnum species or a larger proportion of vascular plants, they can still provide a carbon accumulation function that is crucial for climate change mitigation.
Furthermore, paleoecology helps us to identify the effect of restoration in the long-term context. For example, a high-resolution analysis of testate amoebae, which are very sensitive to hydrological change, can indicate even the subtle effects of the rewetting (Fig. 2; Łuców et al. 2022), enabling us to assess the effectiveness of chosen restoration techniques.
Past environmental conditions that shaped peatland vegetation and the hydrological state were very different from current ecosystem states. This, therefore, exposes a restoration paradox, the scale of which is presumably much greater than we are aware of. It opens questions of how to assess whether an ecosystem should be protected when in most cases long-term data are not considered? How to differentiate the natural succession pathways from anthropogenically induced changes in the past? And where should the restoration be directed if paleoecological data show that the protected ecosystem is the result of a past large-scale anthropogenic degradation process?
Glossary
Wetland – according to the Ramsar Convention wetlands are areas of marsh, fen, peatland or water, whether natural or artificial, permanent or temporary, with water that is static or flowing, fresh, brackish or salt, including areas of marine water the depth of which at low tide does not exceed six meters.
Peatland – a general term that characterizes an area possessing the peat (a layer usually at least 30 cm thick), while mire is a term for wet terrain dominated by living peat-forming plants.
Bog – ombrotrophic peatland dominated by Sphagnum mosses fed by rainwater, highly acidic and isolated from the minerotrophic catchment waters, might be open or wooded.
Fen – minerotrophic peatland that might be open (with e.g. sedges and brown mosses) or wooded, fed by rainwater and ground waters, alkaline and nutrient-rich.
Poor-fen – weakly minerotrophic peatland that is intermediate between fen and bog; usually dominated by Sphagnum mosses, but with hydrology and water chemistry like fens.
Acknowledgements
Authors were supported by the National Science Centre (Poland) (grant nos. 2020/39/D/ST10/00641, 2015/17/B/ST10/01656, 2015/17/B/ST10/03430).
affiliations
1Climate Change Ecology Research Unit, Adam Mickiewicz University, Poznań, Poland
2Max Planck Institute for the Science of Human History, Jena, Germany
3Past Landscape Dynamics Laboratory, Polish Academy of Sciences, Warsaw, Poland
contact
Katarzyna Marcisz: marciszamu.edu.pl
references
Czerwiński S et al. (2021) Quat Sci Rev 259: 106902
Hancock MH et al. (2018) App Veg Sci 21: 167-178
Hennebelle A et al. (2018) For Ecol Manage 430: 157-165
Kołaczek P et al. (2018) Holocene 28: 595-608
Kreyling J et al. (2021) Nat Commun 12: 5693
Lamentowicz M et al. (2020) Sci Rep 10: 18775
Łuców D et al. (2022) Quat Int 607: 65-78
Marcisz K et al. (2015) Quat Sci Rev 112: 138-152
Rydin H, Jeglum JK (2013) The biology of peatlands (Second Edition). Oxford University Press, 398 pp
Sirin A et al. (2021) Land 10: 1200
Słowiński M et al. (2019) J Environ Manage 236: 755-768
Swindles GT et al. (2019) Nat Geosci 12: 922-928
Talbot J et al. (2010) J Veg Sci 21: 143-156
Max Finlayson![]() 1 and Peter Gell
1 and Peter Gell![]() 2
2
Given changes due to the direct drivers of change in wetlands and the absence of suitable reference conditions, we recommend that the Ramsar Convention on Wetlands develops guidance for managers to better understand the nature and trajectory of change in wetlands, and to identify preferred ecological conditions.
The state of wetlands
The Ramsar Convention on Wetlands (see ramsar.org and Gell 2017 for background on the Convention) has reported that national governments are having difficulty in meeting their obligation to maintain the ecological character of internationally important wetlands (Ramsar sites) and other wetlands in their territory (Ramsar Convention on Wetlands 2018; Convention on Wetlands 2021). This indicates a need to better understand the drivers of change in wetlands to inform management decisions, and to identify the trajectories of ecological change.
Key obligations accepted by national governments under the Ramsar Convention are to maintain the ecological character of wetlands they designate as internationally important (known as Ramsar sites), and to make wise use of other wetlands in their territory (Finlayson et al. 2011; Pritchard 2021). These concepts are defined as follows: (1) the ecological character of a wetland is the combination of the ecosystem components, processes, and benefits/services that characterize the wetland at a given point in time; (2) wise use of wetlands is the maintenance of their ecological character, achieved through the implementation of ecosystem approaches, within the context of sustainable development (Finlayson et al. 2011, page 185). Hence, national governments commit to maintaining or restoring wetland ecosystems, their biodiversity, ecological processes, and ecosystem services.
Ecological character of wetlands
The ecological character of wetlands is a consequence of their history, and the ecological trajectories that have been set in place through natural succession and the influence of humans. The former may have occurred over millennia, with more recent human impact arising from pollutants and changes in land and water use within wetlands, and their catchments.
As the maintenance of the ecological character of each Ramsar-listed wetland is judged against an agreed baseline, it is necessary to identify and describe that baseline in sufficient detail for managers to use as a reference, including accommodating natural variability and ecological succession (Pritchard 2021). In some cases, managers have accepted that the ecological character at the time of listing as a Ramsar site was the appropriate baseline. However, many wetlands were in long-term ecological decline long before listing as Ramsar sites, including those documented by Gell et al. (2016). This points to a need to provide information about the nature of change in wetlands, over a range of time-scales, and to improve understanding about their present condition and the importance of anthropogenic drivers. This includes understanding whether paleoecological approaches could be combined with other approaches to understand benchmarks of change, as well as the rates and direction of change, both historic and anthropogenic.
Wetland wise use and long-term change
Guidance for the wise use of wetlands has been developed through the Convention to provide wetland managers with technical knowledge for making decisions for managing and restoring wetlands, including for restoring water regimes and limiting the spread of invasive species. This guidance has not hitherto included how to establish baselines for determining change in ecological character, nor how to respond to variations and change due to climate change (Finlayson 2013). The examples outlined below illustrate how investigations of past change and trajectories can be important for managers.
 |
|
Figure 1: Loch Ruthven has undergone eutrophication, attributed to increased cattle stocking (photo credit: S. Elliot). |
(1) Loch Ruthven, Scotland
Loch Ruthven (Fig. 1) was listed as a Ramsar site in 1996 for its role in maintaining biodiversity in a biogeographic region. Its Ramsar Information Sheet1 (2006) identified it as a mesotrophic lake, and identified its role in supporting breeding pairs of the waterbird, the Slavonian Grebe. Short sediment cores spanning almost 200 years revealed changes in the lake (Brooks et al. 2012). Diatom-inferred total phosphorus revealed the lake to have been oligo-mesotrophic before the 1920s; it is now approaching eutrophic levels. Preserved chironomid (midges) head capsules (a measure frequently used in palaeoecological and palaeoclimate studies) showed that an increase in productivity drove increases in food resources for grebe chicks. This eutrophication trend, attributed to increased cattle stocking, has increased the security of a key bird species, central to the listing of the Ramsar site. Care will be needed to avoid a critical transition that may impact the grebes and other biota.
 |
|
Figure 2: Lake Urmia in Iran is undergoing change in response to drying conditions (photo credit: M. Moser). |
(2) Lake Urmia, Iran
Lake Urmia (Fig. 2), listed as a Ramsar site2 in 1975, was the world's second largest hypersaline lake until 1995 when the water level and area started to decline; it is now almost desiccated (Alizadeh-Choobari et al. 2016). This led to the loss of amenity values (Schmidt et al. 2020), exposure of island refugia for waterbirds, and reduced productivity of Artemia, the main food for migratory flamingos (Schulz et al. 2020). Lake levels have fluctuated greatly with several high lake phases occurring through the Late Holocene (Haghipour et al. 2020); however, the present trend appears to be the result of a drying climate (Schulz et al. 2020; Alizadeh-Choobari et al. 2016) and the use of the catchment's water sources. While the drying of the lake was reported to the Ramsar Secretariat in 2011, such multi-faceted drivers of change challenge restoration efforts owing to the broad bounds of past ecological states revealed through paleoclimate records and the challenge of attributing the change to climate versus human forcing, and local- versus global-scale influences.
 |
|
Figure 3: Ocean entrance to Gippsland Lakes has enabled increased inflow of marine water (photo credit: P. Gell). |
(3) Gippsland Lakes, Australia
The Gippsland Lakes (Fig. 3) in southeast Australia were listed as a Ramsar site in 1982. Its Ramsar Information Sheet3 (1999) reveals changes in water quality attributable to the construction of a permanent opening to the sea in 1889 to allow for navigation and access for ocean fisheries, as well as through farming in the catchment. Hazardous algal blooms have impacted the amenity of the system, yet pigment and isotope analyses from sediment cores show that these were also naturally prevalent prior to opening, owing to nutrient releases under stratified conditions (Cook et al. 2016). In fact, the increased inflow of marine waters acted to suppress blooms owing to higher salinities, at least until the 1940s when nutrient releases from the hinterland increased lake productivity. Recent deepening of the opening has seen an increase of tidal water in the estuary and is resulting in ecological conditions that have hitherto not been previously recorded (Boon et al. 2016), with the death of fringing vegetation and sea-grass, increases in marine biota, and unprecedented cyanobacteria blooms. The system has experienced ongoing change, since well before its listing as a Ramsar site. Future sea-level rise, reduced effective rainfall, the diversion of river flows, and the further intensification of irrigated agriculture may drive the system into novel states that comprise new combinations of species, and are self-sustaining.
Supporting analyses of the trajectories of change in wetlands
These examples raise questions about the adequacy of recent baseline conditions at the time of Ramsar listing for assessing change in the described ecological character. The examples and lessons are highly pertinent to managers, given the expected onset of further change as a consequence of global forces and the inadequacy of many past baselines (see discussion in Kopf et al. 2015). Given that many baseline, or past reference conditions, may not be suitable for future management purposes, it is recommended that the Convention develops guidance to enable managers to better understand the nature and trajectory of change in wetlands and for determining preferred ecological conditions.
affiliations
1School of Biological, Earth and Environmental Sciences, University of New South Wales, Australia
2School of Science, Psychology and Sport, Federation University Australia, Ballarat, Australia
contact
Max Finlayson: colin_maxwell.finlaysonunsw.edu.au
references
Alizadeh-Choobari O et al. (2016) Int J Clim 26: 4276-4286
Boon PI et al. (2016) Mar Freshw Res 67: 721-737
Brooks SJ et al. (2012) J Paleolimnol 47: 631-644
Cook PLM et al. (2016) Biogeosci 13: 3677-3686
Finlayson CM et al. (2011) J Inter Wildl Law Policy 14: 176-198
Finlayson CM (2013) Hydrobio 708: 145-152
Gell PA (2017) PAGES Mag 25: 86-87
Gell PA et al. (2016) Mar Freshw Res 67: 869-879
Haghipour N et al. (2020) EGU General Assembly 2020: EGU2020-7726
Kopf RK et al. (2015) BioSci 65: 798-811
Pritchard D (2021) Mar Freshw Res, doi:10.1071/MF21260
Schmidt M et al (2020) Geojournal 86: 2149-2163
Schulz S et al. (2020) Sci Rep 10: 236
Links
1rsis.ramsar.org/RISapp/files/RISrep/GB855RIS.pdf
2rsis.ramsar.org/RISapp/files/RISrep/IR38RIS.pdf
3rsis.ramsar.org/RISapp/files/RISrep/AU269RIS.pdf
Walter Finsinger![]() 1, E. Cagliero
1, E. Cagliero![]() 1,2, D. Morresi
1,2, D. Morresi![]() 3, L. Paradis
3, L. Paradis![]() 1, M. Čurović
1, M. Čurović![]() 4, M. Garbarino
4, M. Garbarino![]() 3,
3, ![]() 2,
2,![]() 3, V. Spalević
3, V. Spalević![]() 5, E. Lingua
5, E. Lingua![]() 2 and R. Motta
2 and R. Motta![]() 3
3
The long-term history of small and fragmented old-growth forests can provide insights into the legacies of past environmental changes and land-use history in the surrounding landscapes, which can support nature protection and restoration ecology measures.
The value of old-growth forests
Primary and old-growth forests are of great value for sustainable forest management and restoration. By virtue of their structural and compositional complexity, they are highly biodiverse when compared with managed forests in the same ecological region, as they offer a variety of microhabitats such as large pieces of coarse, woody debris, providing shelter to a range of taxa. These naturally regenerated forests of native trees are thought to have developed dynamically for a long period of time without large stand-replacing disturbances, and show no indication of human activities (Barredo et al. 2021). They offer unique opportunities to characterize the effects of natural disturbances and the structural heterogeneity emerging from natural forest dynamics. This is important knowledge to develop close-to-nature forest management practices aimed at emulating natural processes and features in second-growth forests (Schütz et al. 2016).
In Europe, primary and old-growth forests are very rare, generally small, fragmented, and less abundant (< 3% of the total forest area) than on other continents (Sabatini et al. 2018). Currently, these forests are embedded in landscape mosaics bearing variously managed patches that are sometimes strikingly different in species composition and structure. Thus, the small and fragmented ecosystems are threatened along fragment edges by land-use activities (e.g. logging) and anthropogenic disturbances (e.g. fire ignitions) that may initiate long-term changes to the structure of the remaining fragments, thereby hindering the development or the continuity of the old-growth forest stage.
Old-growth forests are typically late-successional forests that contain structures and species that are markedly different from forests of earlier stages. Their distinguishing features include high amounts of deadwood, presence of old trees approaching their natural longevity (which is often much higher than the management rotation cycle for a given tree species), and a patchwork of heterogeneously aged stands arising through small-scale gap dynamics (Barredo et al. 2021).
The current old-growthness is generally assessed using field-based methods, such as dendroecology and forest surveys, and remote sensing (Barredo et al. 2021). However, these methods cannot provide information on the longer-term history of an area. Paleoecology, instead, is sometimes the best, and only, tool for documenting the continuity of forest ecosystems and the legacies of land-use history, climate change, and disturbances (such as fire) on present-day forest composition. This information is critical for clarifying conservation and restoration objectives because mosaic landscapes are often the result of legacy effects arising from complex interactions between natural and human disturbances (Whitlock et al. 2018). In this context, stand-scale paleoecology can be particularly valuable as it allows us to address the history of small and fragmented ecosystems (Bradshaw and Zackrisson 1990; Foster et al. 1996).
Old-growth forests remnants in the Balkans
The causes of the current fragmented distribution of old-growth forests in Europe is a long-standing question in ecology. They are arguably remnants of formerly larger extents of "primeval forests", "virgin forests", "climax forests", or "Urwald" that have been shattered due to major human imprints on forest ecosystems over past millennia (Birks and Tinner 2016). However, primary and old-growth forests are particularly abundant in some regions, such as the Dinaric Alps, possibly due to early protection of forests and lower historical human pressure compared to other mountain ranges in Europe (Sabatini et al. 2018). Indeed, this mountain region is characterized by rugged terrain and land with low agricultural productivity (Kaplan et al. 2009). Nonetheless, detailed records documenting long-term vegetation dynamics, in conjunction with environmental changes, are lacking for the montane zone in this region (Finsinger et al. 2017).
 |
|
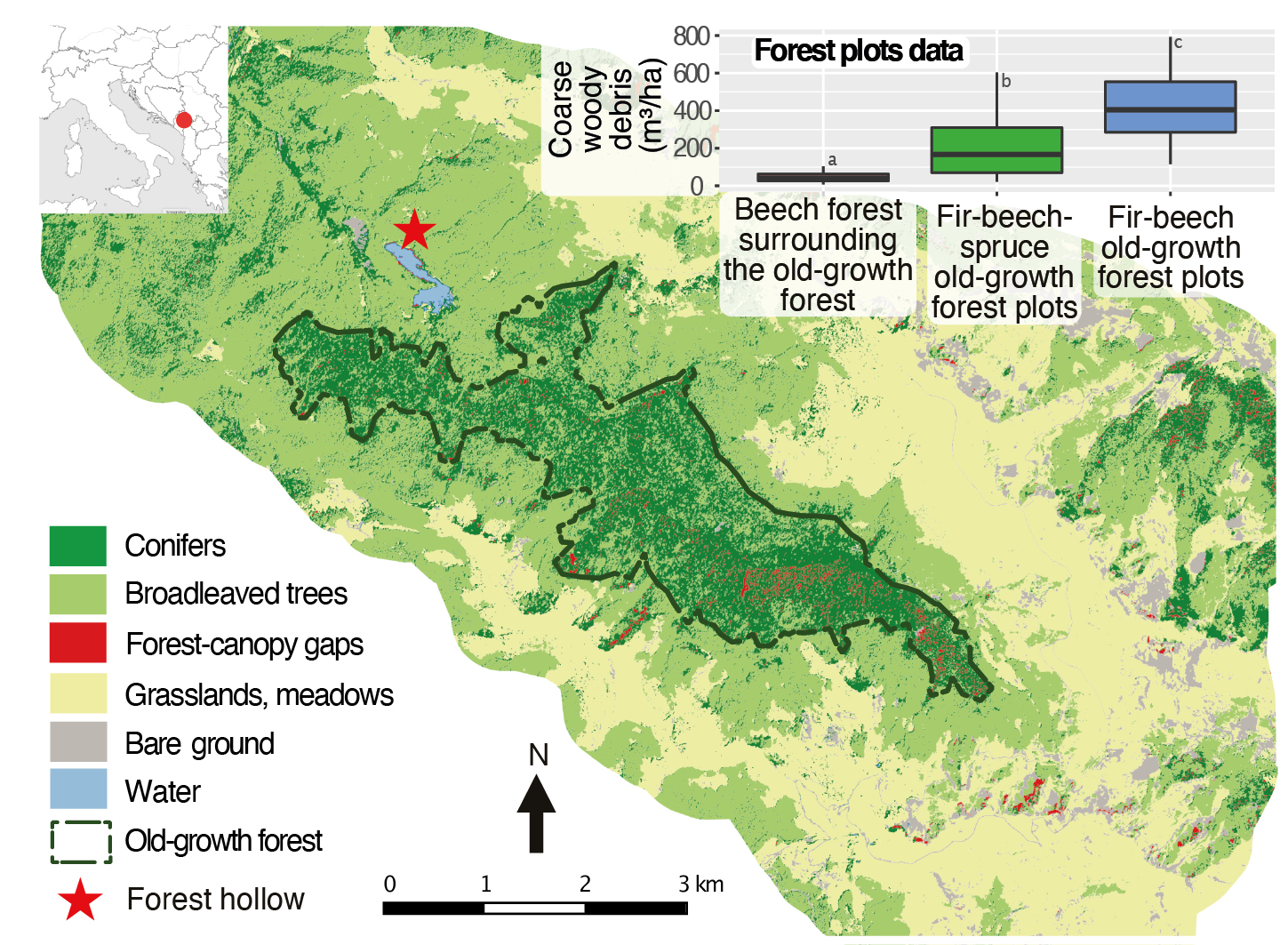
Figure 1: Distribution of land-cover types in the Biogradska Gora valley (Dinaric Alps, Montenegro). The boxplot shows the amount of coarse woody debris inside and outside of the old-growth forest (modified from Cagliero et al. 2021). |
Legacies of past land uses at an old-growth forest's fragment edge
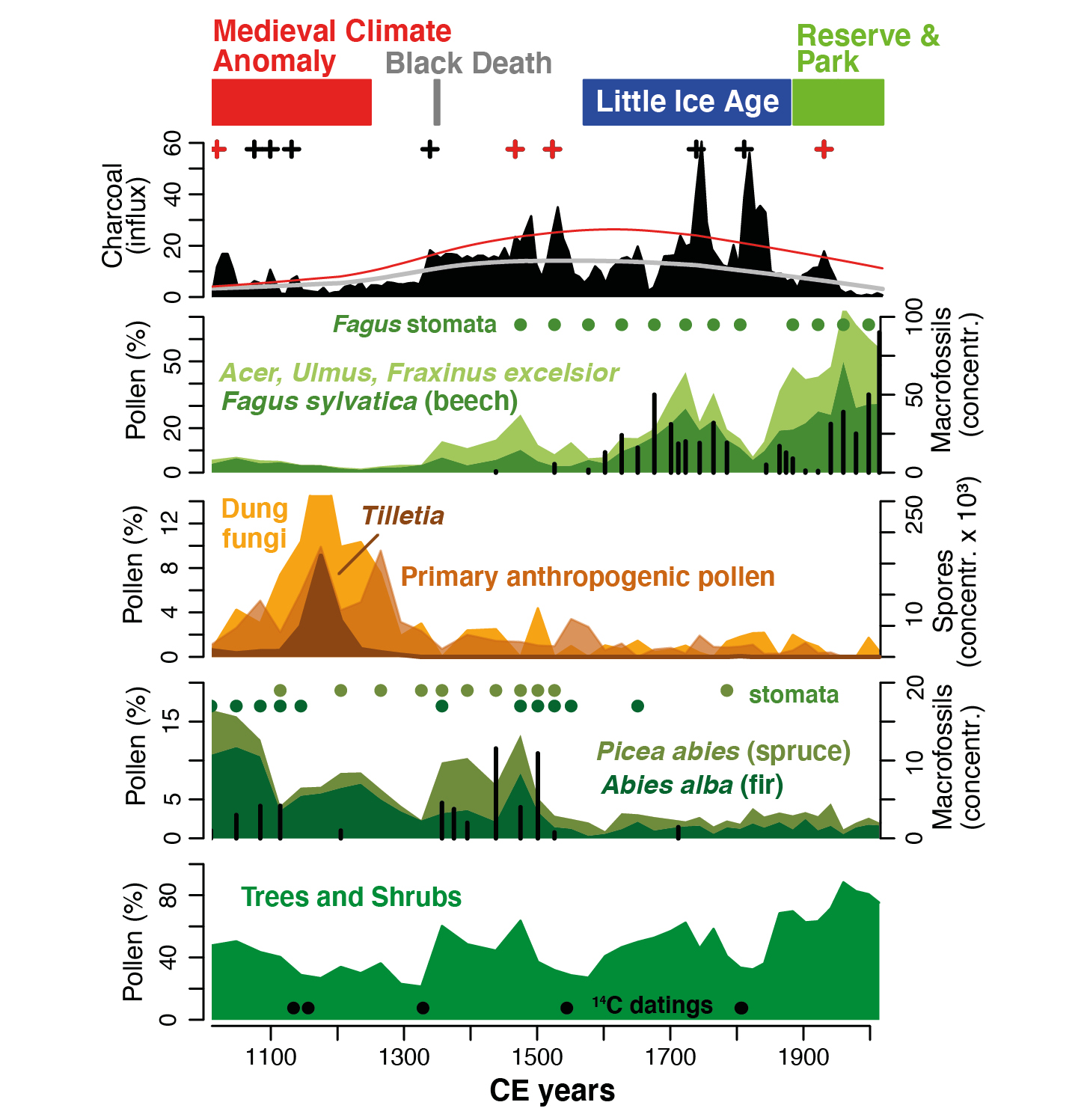
Cagliero et al. (2021) recently presented stand-scale paleoecological records, an assessment of contemporary forest structure, and dendrochronological data from the edge of the Biogradska Gora old-growth forest (Dinaric Alps, Montenegro), one of the largest in Europe. It provides new insights into the long-term dynamics of these fragmented ecosystems. Like other old-growth forests in the montane zone of the Balkans, Biogradska Gora's old-growth forest is dominated by fir (Abies alba) and beech (Fagus sylvatica) with sparse spruce (Picea abies) (Motta et al. 2015). However, unlike many other old-growth forests, this one is surrounded by grasslands and meadows, and by almost pure beech stands. These stands are characterized by small, mostly multi-stemmed trees, the amount of coarse woody debris is negligible (Fig. 1), and charcoal kilns are present. By contrast, the fir-beech-spruce dominated old-growth forest has a mixed and multilayered structure shaped by gaps of different sizes, a large amount of coarse woody debris, and very large and old trees (> 500 years old), indicating that this part of the forest developed dynamically for a long period of time without human and natural stand-replacing disturbances.
The well-dated pollen, spore, stomata, plant-macrofossil, and charcoal records from a small forest hollow at the current edge of the old-growth forest (Fig. 1) document the reduction of fir and spruce during the Middle Ages, when the land was used for cereal crop cultivation and cattle herding, and after local, higher-severity catchment fires occurred (Fig. 2). This evidence supports the notion that historical land-use pressure reduced the extent of old-growth forests.
After intensive local land use ceased (in the mid-14th century, approximately at the time of the Black Death pandemic) and during the cooler Little Ice Age, beech-dominated stands developed in the area surrounding the old-growth (Fig. 2). The legacy of past land use is still visible as the almost pure beech stands show less old-growthness than other European beech-dominated old-growth forests. Conversely, the formal protection of the Biogradska Gora forest as a royal hunting reserve and as a national park since the late 19th century prevented intensive land use, and has strongly reduced biomass burned in recent times, which has allowed for the persistence of the beech stands in the buffer zone. This protected zone may offer a habitat for adaptation to future environmental changes, such as the expansion of the fir old-growth, as fir has more potential than spruce and beech under warmer and drier conditions (Vitasse et al. 2019).
 |
|
Figure 2: Local vegetation and fire-disturbance dynamics at the current edge of Biogradska Gora's old-growth forest as reconstructed with the sediments from a small forest hollow. The charcoal-influx graph includes the long-term trend of charcoal accumulation (grey line) and identified fire episodes (crosses; red crosses: higher-severity catchment fires). Pollen and spores (filled areas), stomata (full circles), and plant macrofossils (black bars) illustrate the vegetation dynamics and land-use history (primary anthropogenic pollen and Tilletia spores: cereal crop cultivation; cattle herding: dung-fungi spores). Full black circles (bottom) show the location of radiocarbon dates (modified from Cagliero et al. 2021). |
Outlook
Old-growth forests have captured the attention of foresters and conservationists alike (Fröhlich 1930). However, there are still substantial conservation and restoration gaps, as primary and old-growth forests are only partially representative of the full range of European forest types (Sabatini et al. 2018). Paleoecology, in conjunction with structural and dendrochronological research, can unfold the history of Europe's primary and old-growth forests. Thereby, their responses to past environmental changes, as well as legacy effects of past land use and of disturbances in surrounding landscapes, can be assessed. This may be important as strategies for the preservation and restoration of old-growth forests should acknowledge past environmental changes, including land-use history, and anticipate future environmental changes.
Although primary and old-growth forests are generally included in protected areas (Sabatini et al. 2018), they may be vulnerable to climate change and associated changing disturbance regimes (wind, pathogens, or fire; Seidl et al. 2017). For instance, several of the smaller old-growth forests that bear fire-sensitive species such as Abies alba may be at risk if fire-frequency becomes excessive. In this context, identifying and protecting primary and old-growth areas and their buffer zones, as well as protecting and restoring secondary forests that may represent future primary and old-growth ecosystems (Barredo et al. 2021; EU's Biodiversity Strategy for 2030: ec.europa.eu/environment/strategy/biodiversity-strategy-2030_en) could be of paramount importance to promote native forests. Such actions may likely increase ecosystem resilience to future climate change and be helpful to anticipate environmental changes (Henne et al. 2015).
affiliations
1ISEM, Univ Montpellier, CNRS, EPHE, IRD, France
2Department of Land, Environment, Agriculture and Forestry, University of Padova, Legnaro, Italy
3Department of Agricultural, Forest and Food Sciences, University of Torino, Grugliasco, Italy
4Biotechnical Faculty, University of Montenegro, Podgorica, Montenegro
5Geography Department, University of Montenegro, Nikšić, Montenegro
contact
Walter Finsinger: walter.finsingerumontpellier.fr
references
Bradshaw RHW, Zackrisson O (1990) J Veg Sci 1: 519-528
Cagliero E et al. (2021) Veg Hist Archaeobot, doi:10.1007/s00334-021-00862-x
Finsinger W et al. (2017) Quat Sci Rev 167: 63-77
Foster DR et al. (1996) Trends Ecol Evol 11: 419-424
Fröhlich J (1930) Centr Gesamte Forstwesen 56: 49-68
Henne PD et al. (2015) Front Ecol Environ 13: 356-362
Kaplan JO et al. (2009) Quat Sci Rev 28: 3016-3034
Motta R et al. (2015) Plant Biosyst 149: 966-975
Sabatini FM et al. (2018) Divers Distrib 24: 1426-1439
Schütz J-P et al. (2016) Ann For Sci 73: 911-921
Seidl R et al. (2017) Nat Clim Chang 7: 395-402
César Morales-Molino![]() and Christoph Schwörer
and Christoph Schwörer![]()
Drawing on several case studies from the Iberian Peninsula, we show how specifically designed paleoecological research can be a useful tool to guide ecosystem management, conservation, and restoration.
The Mediterranean region is well known for its outstanding diversity of species and ecosystems. This natural heritage is, nevertheless, threatened not only by ongoing climatic change, but also by legacies of multi-millennial land use and shifts in land-use patterns, which can significantly alter natural disturbance regimes. This raises questions for management and restoration of ecosystems that can be informed by long-term data from paleoecology. Here, we discuss three examples from the Iberian Peninsula that relate to fire management, herbivore management, and the conservation of cultural landscapes.
Changing fire regimes and their impact on Pinus nigra forests
Depopulation of rural areas and abandonment of traditional farming practices are currently leading to increased woodland encroachment, and subsequent accumulation of dead biomass (Keeley et al. 2012). Under particularly favorable conditions for fire spread and the presence of (usually anthropogenic) ignition sources, these landscapes offer the perfect setting for catastrophic fires. Concerned by this relevant and increasingly pressing issue, ecosystem managers often try to adapt forest management strategies to prevent these large and severe fires, or at least limit their spread.
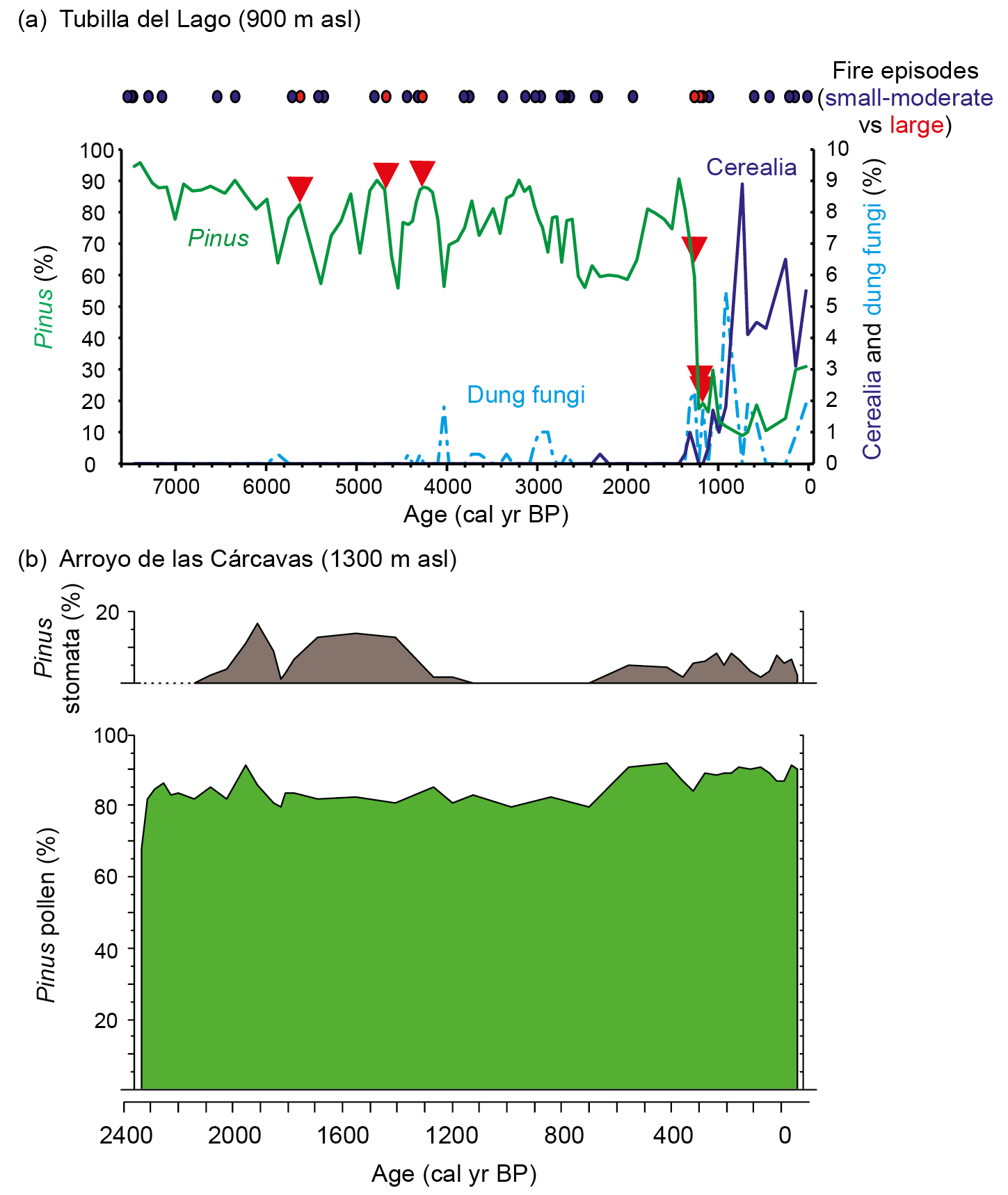
In the Northern Iberian Plateau (central-northern Spain), we investigated the causes of the regional demise of Pinus nigra forests during historical times using a multi-proxy approach including pollen, conifer stomata, dung fungal spores, and charcoal particles, alongside peak detection analysis (Morales-Molino et al. 2017a). The results show that pine forests were the dominant vegetation in the region for most of the Holocene, under a mixed fire regime consisting of frequent low-severity (ground) fires and rare high-severity (crown) fires (Fig. 1). Consistent with its life-history traits (Tapias et al. 2004), P. nigra was able to resist ground fires but, unexpectedly, stands could also recover after single crown fires despite lacking specific adaptations (e.g. serotinous cones). However, when crown fires became very frequent with the intensification of arable and pastoral farming during the Middle Ages (1200–1000 yr cal BP), pine forests rapidly declined and even disappeared regionally (Fig. 1a).
 |
|
Figure 1: (A) Paleoecological record from Tubilla del Lago (central-northern Spain). Top: Fire episodes as reconstructed using peak detection analysis (Higuera et al. 2009) applied to the high-resolution macroscopic charcoal series, distinguishing between small-moderate ("surface fires") and large ("crown fires") peaks. Bottom: Abundances of Pinus pollen and the main local indicators of farming activities (Cerealia type, dung fungal spores). Red triangles denote the occurrence of crown fires. Modified from Morales-Molino et al. (2017a); (B) Pollen and stomata evidence of the dominance of Pinus in the Sierra de Guadarrama for the past > 2000 yr. Modified from Morales-Molino et al. (2017b). |
The take-home message for forest managers who want to preserve the species-rich Pinus nigra forests currently thriving in the circum-Mediterranean mountains is that fuel loads need to be managed using prescribed ground fires, in order to prevent high-severity crown fires. This low-severity fire regime will, in turn, also promote seedling recruitment.
Unprecedented herbivore densities and the fate of Mediterranean mires
Another worrying aspect of ongoing land abandonment, in combination with the local extirpation of natural predators, is the dramatic increase in the densities of wild ungulates that prevents the recruitment of palatable woody species and causes potentially damaging disturbance when large herds visit wetlands (e.g. mires, springs, ponds) during the dry Mediterranean summers. Managers of protected areas need scientific knowledge about the carrying capacity of these ecosystems, i.e. the herbivore densities that the ecosystem can tolerate without risking its functionality.
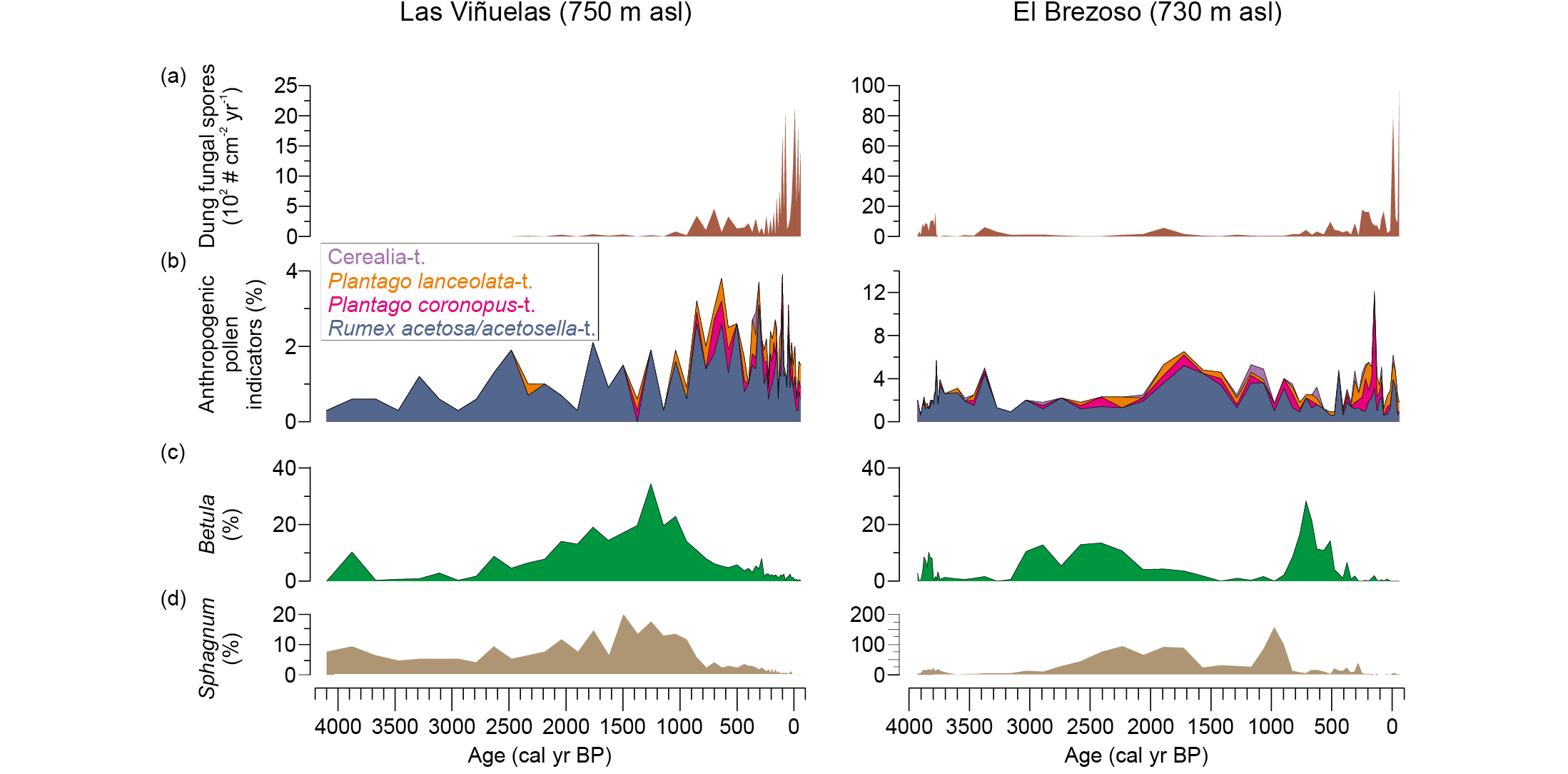
In the Cabañeros National Park (central-southern Spain), current densities of wild ungulates (e.g. red deer, wild boar) are so high that mires are regularly damaged, and tree regeneration is seriously affected. We conducted paleoecological research on two peat sequences to investigate the past impact of herbivore densities (wild ungulates and livestock), inferred from dung fungal spores (Baker et al. 2016), and land use on the mires of this protected area (Morales-Molino et al. 2019). The pollen and dung fungal spore data show that locally growing Betula stands, as well as Sphagnum mats, declined and even went extinct when pastoral farming increased during the Middle Ages (1000–800 yr cal BP; Fig. 2). The influxes of dung fungal spores suggest that the extremely high current herbivore densities are unprecedented in the context of the past 4000 years. Therefore, herbivores pose a severe threat to extant mires in the region, including those where relict birch stands still survive. We strongly recommend fencing the mires to prevent further destruction by ungulates, as well as controlling the populations of these herbivores, preferably by the re-introduction of natural predators such as wolf or Iberian lynx, which went regionally extinct just a few decades ago.
 |
|
Figure 2: Paleoecological sequences from the Cabañeros National Park (central-southern Spain). Abundances of (A) dung fungal spores, a proxy for past herbivore density (Baker et al. 2016); (B) anthropogenic pollen indicators, directly or indirectly related to farming activities and disturbance; (C) Betula pollen; (D) Sphagnum spores, which are related to the extent and integrity of the mires. Modified from Morales-Molino et al. (2019). |
What is "natural" in the Mediterranean region?
The millennial history of farming and forest use in the Mediterranean region makes it hard to assess the naturalness of certain ecosystems often considered as purely anthropogenic based on a shorter-term perspective. Precise knowledge about the "natural" composition and structure of vegetation is of paramount importance if the goal is to restore ecosystems to their "original" state, preceding the onset of heavy human impact.
The question of what the natural vegetation in an area would be remains crucial in many Mediterranean areas that were intensively disturbed by land use in the past. The "potential natural vegetation" concept, i.e. mature vegetation in the absence of human impact, has been widely used during the past few decades in this context, despite being subject to debate and controversy (e.g. Carrión and Fernández 2009; Chiarucci et al. 2010; Farris et al. 2010; Loidi and Fernández-González 2012). The long-term empirical perspective of paleoecology on vegetation dynamics, in particular in response to human impacts, allows a largely refined discussion of this topic (e.g. Carrión and Fernández 2009).
In the Valsaín forests of the Sierra de Guadarrama National Park, in the Iberian Central Range, the potential natural vegetation would consist of Quercus pyrenaica forests, whereas pine forests are thought to have been favored by humans (Morales-Molino et al. 2017b). We analyzed several mires using a multi-proxy approach (pollen, conifer stomata, plant macrofossils) to investigate whether Pinus sylvestris forests occur naturally at relatively low elevation in these mountains. The high pollen percentages along with the finds of Pinus stomata and macrofossils demonstrate that Scots pine has dominated forests as low as 1300 m asl for at least the past 2500 years (Fig. 1b). Paleoecological evidence therefore supports the naturalness of pine forests and refutes the need to favor Quercus pyrenaica in forest management.
Conclusions and outlook
The three case studies presented above illustrate how paleoecological data provide relevant information to guide the conservation and restoration of Mediterranean ecosystems. The first two examples show how paleoecological records can assist in restoring fire regimes and herbivore densities that maintain and recover vegetation. The third example shows how the long-term perspective provided by paleoecology, particularly with respect to past human impacts on vegetation dynamics, can help resolve some of the controversies raised by the "potential natural vegetation" concept (e.g. Carrión and Fernández 2009). However, we acknowledge the need for refinements and novel techniques for a broader use of these data in a global change scenario.
For instance, dynamic vegetation models allow us to disentangle the past roles of climate, disturbance, and human activities in driving vegetation changes, improving predictions of future vegetation dynamics under different scenarios of climate change and land use (e.g. Henne et al. 2015; Schwörer et al. 2014). Additionally, population genetic analyses on ancient DNA preserved in plant macrofossils may provide unique information on which variant of a given tree species would be best suited to revive a population that became locally extinct in the past (Schmid et al. 2017).
affiliation
Institute of Plant Sciences and Oeschger Centre for Climate Change Research, University of Bern, Switzerland
contact
César Morales-Molino: cesar.moralesips.unibe.ch; cesarmoralesdelmolinogmail.com
references
Baker AG et al. (2016) Methods Ecol Evol 7: 1273-1281
Carrión JS, Fernández S (2009) J Biogeog 36: 2202-2203
Chiarucci A et al. (2010) J Veg Sci 21: 1172-1178
Farris E et al. (2010) J Biogeog 37: 2211-2213
Henne PD et al. (2015) Front Ecol Environ 13: 356-362
Higuera PE et al. (2009) Ecol Monogr 79: 201-219
Loidi J, Fernández-González F (2012) J Veg Sci 23: 596-604
Morales-Molino C et al. (2017a) J Ecol 105: 634-646
Morales-Molino C et al. (2017b) Glob Planet Change 152: 64-75
Morales-Molino C et al. (2019) Ecology 100: e02833
Perea R, Gil L (2014) For Ecol Manag 312: 47-54
Schmid S et al. (2017) Methods Ecol Evol 8: 1374-1388
Sandra Nogué![]() 1,2, L. de Nascimento
1,2, L. de Nascimento![]() 3, W.D. Gosling
3, W.D. Gosling![]() 4, N.J.D. Loughlin
4, N.J.D. Loughlin![]() 5, E. Montoya
5, E. Montoya![]() 6 and J.M. Wilmshurst
6 and J.M. Wilmshurst![]() 7,8
7,8
Recent work within restoration ecology has highlighted the importance of incorporating ecological history. Using three complementary examples from New Zealand offshore islands, the tropical Andes, and the Canary Islands, we discuss how restoration goals may be addressed using multiple baselines, or reference conditions, from long-term data.
The idea of using long-term ecological data to measure changes in biodiversity and to improve the effectiveness of conservation strategies has been widely discussed (Willis et al. 2010; Nogué et al. 2017). Importantly, the use of baselines, or reference conditions, has already been recognized in environmental policy such as in the conceptual framework for the 2019 Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services (IPBES) global assessment report, and the United Nation decade on ecosystem restoration (2021–2030). It is, therefore, clear that there is now an urgent need to organize and standardize baselines if we want to implement successful restoration actions taking into account the complexity of managing both ecological and cultural systems (Jackson and Hobbs 2009; Nogué et al. 2021). We highlight three case studies to show how restoration goals may be addressed using paleoecologically informed baselines.
(1) Vegetation baselines for New Zealand offshore islands
New Zealand's offshore and outlying islands are important foci for biodiversity protection and conservation, with many supporting species that have been extirpated from the main islands (Bellingham et al. 2010). Most islands have been modified since initial human settlement began ca. 780 years ago, mainly by forest clearance and introduced mammalian pests (particularly rodents, cats, goats and pigs). Conservation efforts in the last 50 years have focused on eradicating alien species and replanting and restoring the "natural" vegetation. However, baselines have usually been informed by historic descriptions of vegetation and/or the composition of nearby forested locations.
Paleoecological records are recognized in New Zealand as an additional tool to document natural pre-human vegetation baselines and to help guide restoration plans (Wilmshurst et al. this issue). Long-term records have also been combined with local Māori (New Zealand's indigenous Polynesian population) knowledge, or mātauranga, to provide cultural vegetation baselines and to help inform biocultural approaches to island conservation (Lyver et al. 2015). For example, pollen and charcoal records showed how some forested ecosystems were cleared for Māori gardens, and local Māori communities now desire some forest to be restored to states that would provide a range of future benefits to their cultural, social, economic, and ecological needs (Lyver et al. 2015).
 |
|
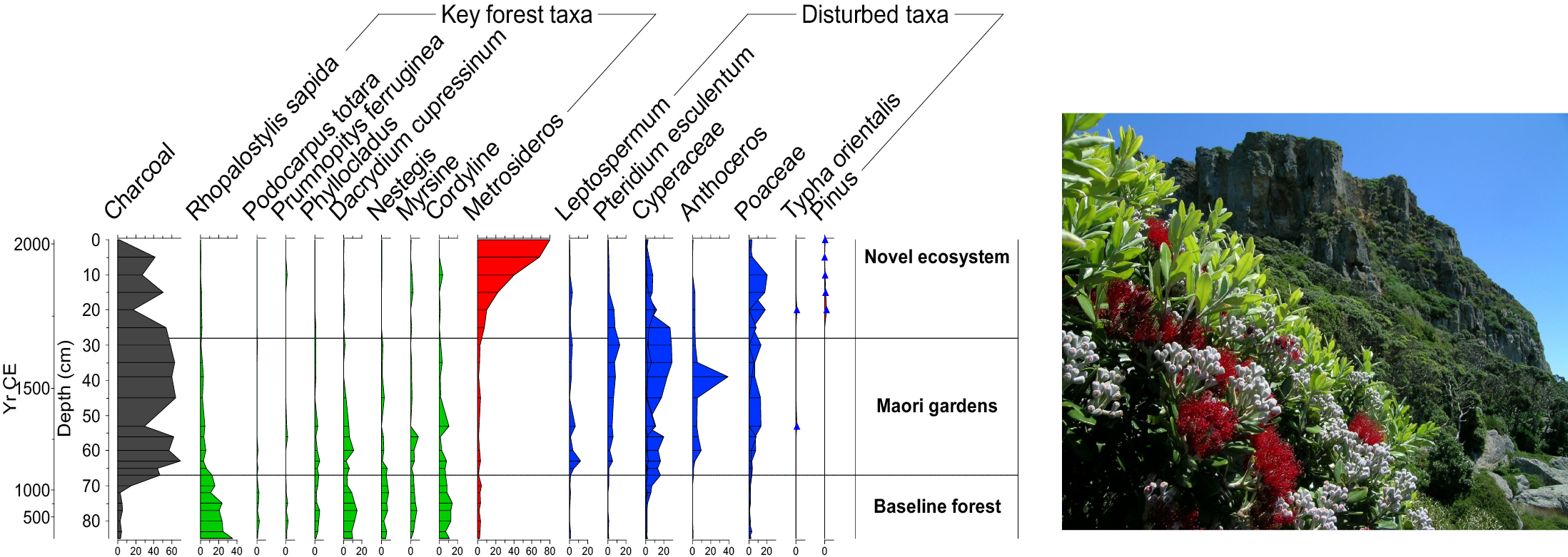
Figure 1: Example from Tawhiti Rahi Island of the Poor Knights Islands (Wilmshurst et al. 2014). Pollen data shows that the current vegetation composition is completely different from ancient ecosystems (photo credit: J.M. Wilmshurst). |
Sometimes paleoecological records can reveal surprises that overturn current ecological understanding. For example, the Poor Knights Islands in northeastern New Zealand are currently covered in a tall forest of native angiosperm trees dominated by Metrosideros. This vegetation composition has long been considered to be in a natural baseline state and is used to inform replanting of degraded islands in the region. However, a 2000 year record of pollen, charcoal and ancient plant DNA from a soil core on the island (Fig. 1) tells a different story (Wilmshurst et al. 2014). The forest was cleared by fire in the 13th century and gardened by Māori for ca. 550 years, followed by 180 years of forest succession after they ceased living there. The pre-human vegetation was completely different to today's forest, consisting of a diverse, conifer-dominated forest (e.g. Dacrydium cupressinum and Prumnopitys ferruginea) with an understory palm (Rhopalostylis sapida) and several other locally extinct tree taxa. Not only was Metrosideros unexpectedly uncommon in the past, but the pre-human baseline has no current analog on any northern New Zealand islands. This, and other paleoecological records from the region, demonstrate the power of the past to help inform future directions for conservation management.
(2) Examples from the tropical Andes
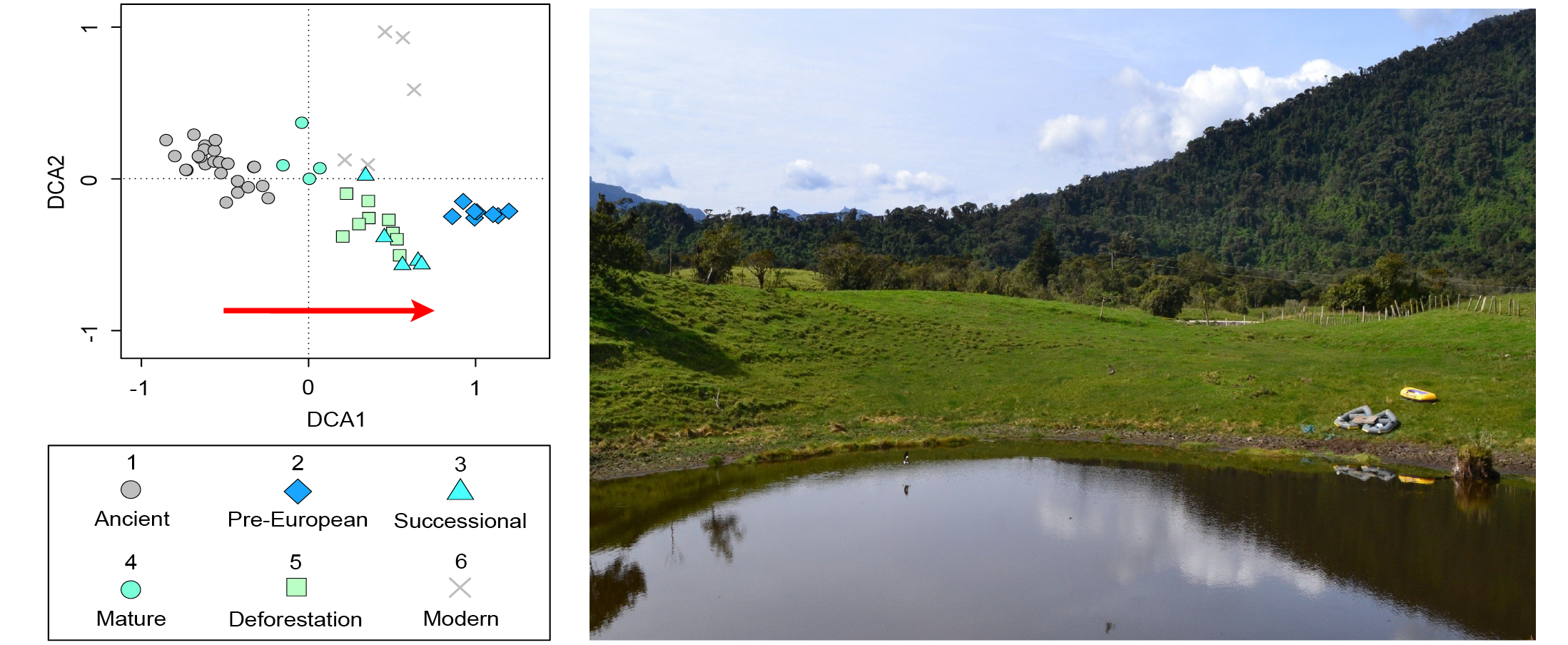
The Andean flank region has particularly caught the attention of researchers and policymakers due to its ecological value, and both its historical and current anthropogenic pressures (Cuesta et al. 2019). Paleoecological studies carried out in the tropical Andes have focused on using past environmental information to anticipate future scenarios of global change, including: (1) observing the synchronicity between environmental change and societal restructuring and adaptation in pre-Columbian populations (Gosling and Williams 2013); (2) pinpointing potential microrefugia locations based on the past dynamics of the Andean flag taxon Polylepis sp. (Valencia et al. 2016); and (3) highlighting shifted historical baselines such as those observed in the 19th century in locations with large pre-Columbian anthropogenic pressure that suffered depopulation following European arrival (Loughlin et al. 2018).
For example, fossil pollen showed a succession of Andean forest driven by changes in land-use. This forest succession was divided into five different vegetation baselines (Loughlin et al. 2018): pre-European (pre-CE 1588 indigenous occupation), successional (CE 1588–1718 European arrival/re-colonization), mature (CE 1718–1819 diminished population), deforestation (post CE 1819, re-colonization), and modern (industrial agriculture). These results were compared to a close site indicative of pre-human arrival vegetation (42 cal kyr BP; ancient), and showed the closest similarity with the post-abandoned mature state (Fig. 2). These new insights from sedimentary cores spanning the past three to four centuries show the potential to characterize meaningful restoration targets and reference ecosystems from paleoecological data (see Rull, this issue).
 |
|
Figure 2: Detrended correspondence analysis (DCA) of pollen data from Andean montane forest grouped into time bins. Red arrow indicates an increasing signal of human impact. Replotted from Loughlin et al. (2018) (photo credit: N.J.D. Loughlin). |
(3) The thermophilous forest of the Canary Islands
The Canarian thermophilous woodland is among the most threatened ecosystems in the Canary Islands (del Arco et al. 2010). The remnants of these woodlands are currently occupying less than 10% of their original area and are undergoing several restoration actions. Describing baselines for these woodlands have proved challenging and is limited to the available ecological information from the few woodlands remaining on the islands.
On the island of Gran Canaria, there have been several restoration programs mostly focused on laurel forest remnants, since only 1% of its original distribution remains on the island today. One program included restoring the Laguna de Valleseco as part of the LIFE Laurisilva XXI project, with a key objective devoted to education (Cárdenes 1998). When this project was implemented, the pre-human baseline for the site was thought to be laurel forest with the presence of e.g. Ilex canariensis, Erica arborea, and Morella faya agreeing with the potential vegetation maps that predicted its presence in the area (del Arco et al. 2006). However, a paleoecological record carried out at the same site suggested a wider distribution of the thermophilous forest in the past, including this northern area. Fossil pollen records showed the unexpected presence of thermophilous species such as Juniperus and the date palm (Phoenix canariensis) until about 2300 years ago when there was an increase in fires, and grasses and shrubs spread over the area (de Nascimento et al. 2016). Before these new insights from the pollen record, Juniperus turbinata was considered a rare tree on the island (del Arco et al. 2006). This mismatch between data sources highlights the utility of combining paleoecological data into restoration planning, and improving understanding around past composition and occurrence of plant communities.
In Teno, on the island of Tenerife (European Union LIFE Habitat project), the reference condition was obtained by a combination of sources, including inventories of locally remaining Juniperus turbinata trees and patches of Juniper woodlands remaining in the archipelago (Tenerife and La Gomera), potential vegetation maps, toponymy (e.g. place name or geographic name), and historical and recent (elderly locals) accounts, to confirm past occurrence of the woodlands (Fernández-Palacios et al. 2008). Ten years after plantation, populations of J. turbinata are showing positive growth trends, with eight tree and shrub species surviving, growing, and some even showing signs of reproduction (flowers and fruits) (Rota et al. 2021).
Conclusions
With these three examples, covering ecosystems varying from high-mountains to islands, and from both hemispheres, we show the importance of incorporating paleoecological knowledge and other sources of long-term data in restoration programs. There are numerous examples of paleoecological research that have highlighted a priori misconceptions on the definition of the natural vegetation (Bush et al. 2014), and the ability of past environmental change data to characterize landscapes that are different to those of today (Rull et al. 2017). Paleo-informed baselines may provide a useful perspective on the variability of ancient systems, the magnitude of biodiversity change, novel ecosystems, and responses to successive waves of cultural transformation.
Acknowledgements
This work was supported by the Spanish Ministry of Science and Innovation (CGL 2009-10939), the Open University through a scholarship to N.J.D.L. (NE/L501888/1), and a NERC fellowship to E.M. (NE/J018562/1).
affiliations
1CREAF, Centre de Recerca Ecològica i Aplicacions Forestals, Bellaterra (Cerdanyola del Vallès), Catalonia, Spain
2School of Geography and Environmental Science, University of Southampton, UK
3Island Ecology and Biogeography Group, Instituto Universitario de Enfermedades Tropicales y Salud Pública de Canarias (IUETSPC), Universidad de La Laguna, Canary Islands, Spain
4Department of Ecosystem and Landscape Dynamics, Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, The Netherlands
5Geography & Environmental Sciences, Northumbria University, Newcastle Upon Tyne, UK
6Laboratory of Paleoecology, Geosciences Barcelona (CSIC), Barcelona, Spain
7Long-term Ecology Laboratory, Manaaki Whenua-Landcare Research, New Zealand
8School of Environment, University of Auckland, New Zealand
contact
Sandra Nogué: s.noguecreaf.uab.cat
references
Bellingham PJ (2010) N Z J Ecol 34: 115-136
Bush MB et al. (2014) Restor Ecol 22: 296-298
Cárdenes JL (1998) Medio Ambiente Canarias 11: 15-16
Cuesta F et al. (2019) Reg Environ 19: 1623-1642
del Arco MJ et al. (Ed) (2006) Mapa de Vegetación de Canarias. GRAFCAN, 552 pp
del Arco MJ et al. (2010) Biodivers Conserv 19: 3089-3140
de Nascimento L et al. (2016) Holocene 26: 113-125
Fernández-Palacios JM et al. (2008) Los bosques termófilos de Canarias (Proyecto LIFE04/NAT/ES/000064). Santa Cruz de Tenerife: Cabildo Insular de Tenerife
Gosling WD, Williams JJ (2013) Holocene 23: 1619-1624
Jackson ST, Hobbs RJ (2009) Science 325: 567-569
Loughlin NJD et al. (2018) Nat Ecol Evol 2: 1233-1236
Lyver POB et al. (2015) Hum Ecol 43: 681-695
Nogué S et al. (2021) Science 372: 488-491
Nogué S et al. (2017) Nat Ecol Evol 1: 0181
Rota F et al. (2021) Scientia Insularum 4: 27-44
Rull V et al. (2017) PAGES Mag 25: 82-83
Valencia BG et al. (2016) New Phytol 212: 510-522
Lindsey Gillson![]() 1, C. Whitlock2, P. Gell
1, C. Whitlock2, P. Gell![]() 3, S. Prader1 and W. Tinner
3, S. Prader1 and W. Tinner![]() 4
4
2021 marked the beginning of the United Nations Decade on restoration ecology. Restoration of ecosystems is essential in slowing biodiversity loss and associated erosion of ecosystem services. However, defining restoration goals in an uncertain and changing world raises fundamental questions of what we are restoring and why. The purpose of this special issue is to explore the contributions of paleoecology in addressing these questions and to encourage better integration of paleoecology into restoration ecology and conservation planning.
What are we restoring?
The most obvious—and yet still underutilized—use of paleoecological data in restoration ecology is to provide reference conditions, especially in ecosystems that have experienced significant anthropogenic degradation over periods of time that extend beyond living memory or historical records (e.g. Finlayson and Gell p. 10; Marcisz et al. p. 12; Hapsari et al. p. 14). Paleoecological data sometimes reveal surprises regarding the extent and composition of vegetation in the past, showing that current vegetation is in fact far from natural, and confirming or rejecting the status of alien species (Nogué et al. p. 4; Wilmshurst and Wood p. 26). Although "naturalness" is a contested term, areas with minimal or light human impact are, nevertheless, an important landscape component in many regions (e.g. Nanavati et al. p. 22; Morales-Molino and Schwörer p. 6; Rull p. 18; Finsinger et al. p. 8). Restoration of desired cultural landscapes can also have benefits to both biodiversity and people (see Rull p. 18).
 |
|
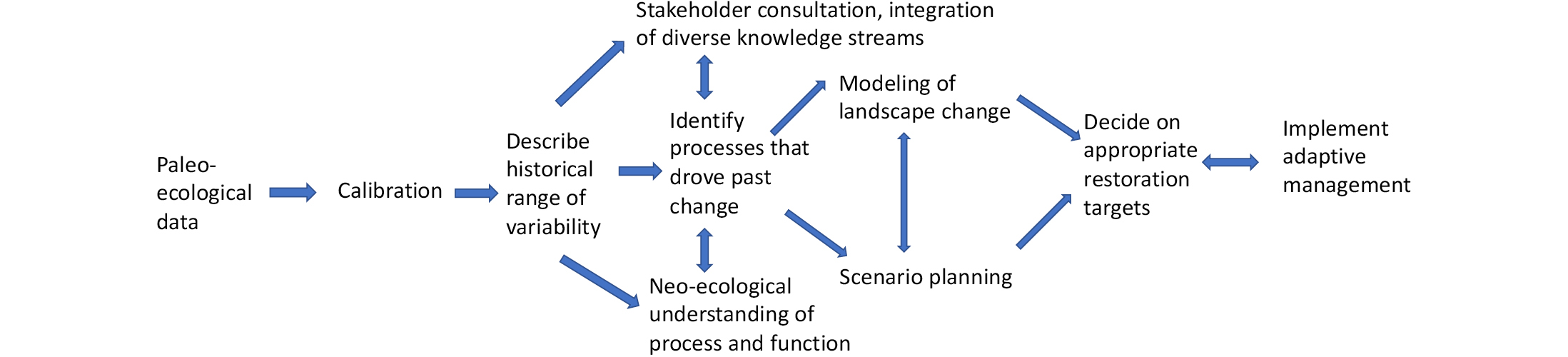
Figure 1: A suggested framework by which paleoecology could be integrated with other disciplines and knowledge streams in a process-based approach to restoration ecology that includes science, modeling, stakeholder consultation and adaptive management. |
Even without significant human impact, most landscapes are dynamic and respond to multiple interacting environmental drivers, including changes in climate, disturbance, land use, and biotic interactions. Understanding the long-term importance of fire and herbivory, for example, is an important scientific contribution from paleoecology, particularly as it relates to climate extremes, land abandonment, and rewilding (Higuera et al. p. 30; Morales-Molino and Schwörer p. 6). Interactions among environmental, biotic, and anthropogenic drivers can also cause shifts between alternate stable states. This is especially likely at ecotones (vegetational transitions), which are sensitive to subtle changes in climate, fire, and land use and therefore present particular challenges for restoration (Nanavati et al.p. 22; Giesecke et al. p. 24).
Why are we restoring? From static "baselines" to dynamic processes
In today's changing world, no-analog climate conditions are increasingly likely in the coming decades, and a return to "natural" conditions may be impossible or undesirable. As a result, there has been a shift in restoration ecology towards a broader range of conservation objectives that considers the degree of past anthropogenic modification, as well as the desired ecosystem function or condition (Chambers p. 16; Rull p. 18). Considerations include which ecosystems will be most vulnerable to future climate and land-use change and which should be prioritised for restoration and conservation actions (Adeleye et al. p. 28; Higuera et al. p. 30). Paleoecology can also guide efforts to maintain critical ecological functions, such as pollination, by revealing unsuspected past interactions in species whose ranges are currently disjunct (Wilmshurst and Wood p. 26). The integration of paleoecology into an inclusive, process-based approach to restoration ecology is illustrated in Figure 1. Note that as the future is uncertain, the implementation approach needs to be adaptive.
Conclusions and ways forward
The papers in this issue demonstrate a huge and largely untapped synergy between the disciplines of paleoecology and restoration ecology. Ensuring that this potential is realized will require a concerted effort by the paleoecological community in seven main areas:
- Better calibration of paleoecological datasets to increase their usefulness as reference frameworks for conservation planning.
- Wider incorporation of new techniques, such as ancient DNA/sedimentary DNA, to document past changes in biodiversity (e.g. see Wilmshurst and Wood p. 26).
- Communicating paleoecological findings in an applied context, wherever possible, so that the information is accessible and available to the restoration ecology community and beyond.
- Greater integration of paleoecology with other disciplines and knowledge streams, including traditional ecological knowledge (see Gil-Romera et al. p. 20 and the special section in this issue "Socio-ecological approaches to conservation" p. 33).
- Showcasing the relevance of historical perspectives in process-based thinking and modeling efforts that guide adaptive management planning for emerging conditions and societal preferences (e.g. Morales-Molino and Schwörer p. 6).
- Validating dynamic modeling outcomes, for example, by comparing sedimentary proxy data with simulations of ecosystem changes in response to climate change, disturbance, and land use.
- Encouraging managers and policy makers to think on time scales longer than a few decades so that paleoecological information becomes routinely incorporated into landscape conservation planning (e.g. the Ramsar Convention; see Finlayson and Gell p. 10; Hapsari et al. p. 14; Chambers p. 16).
The paleoecological community has a vital challenge ahead: that of seamlessly integrating paleoecology and neo-ecology, thereby enabling the mainstreaming of paleoecology into restoration ecology and biodiversity conservation.
affiliationS
1Plant Conservation Unit, Department of Biological Sciences, University of Cape Town, South Africa
2Department of Earth Sciences, University of Montana, Bozeman, USA
3School of Science, Psychology and Sport, Federation University Australia, Ballarat, Australia
4Institute of Plant Sciences and Oeschger Centre for Climate Change Research, University of Bern, Switzerland
contact
Lindsey Gillson: lindsey.gillsonuct.ac.za
Nils Weitzel![]() 1, C. Brierley
1, C. Brierley![]() 2, J. Bühler
2, J. Bühler![]() 1, M. Chevalier
1, M. Chevalier![]() 3, B. Ellerhoff
3, B. Ellerhoff![]() 1, V. Skiba
1, V. Skiba![]() 4 and K. Rehfeld
4 and K. Rehfeld![]() 1,5
1,5
Heidelberg, Germany, and online, 5-7 July 2021
A key question of the paleoclimate community is how paleoclimate data can be used to evaluate long-term predictability in climate models. How can we improve estimates of past climate variability and our understanding of the state and timescale dependency of Earth's climate? "Ping pong" serves as a metaphor to describe the back-and-forth in comparing paleoclimate data with model simulations. This is a core challenge in climate research, which requires a better understanding of proxies as well as the consequences of neglected or poorly simulated processes in climate models.
To address this question, this Climate Variability Across Scales (CVAS; pastglobalchanges.org/cvas) workshop (pastglobalchanges.org/calendar/26970) brought together a diverse pool of ∼60 scientists, ranging from early-career scientists to experienced experts from various fields and different working groups, including CVAS, Speleothem Isotopes Synthesis and AnaLysis (SISAL; pastglobalchanges.org/sisal), 2k Network (pastglobalchanges.org/2k), and the PAGES-endorsed Paleoclimate Modelling Intercomparison Project (PMIP; pmip.lsce.ipsl.fr). One half participated online and the other half gathered at the "Internationales Wissenschaftsforum" in Heidelberg, Germany. Keynote talks focused on climate variability on different temporal and spatial scales, best practices for the joint use of models and proxies, the role of paleoclimate in future predictions, and the state of the art in the analysis and interpretation of various proxy types. Discussions resulting from these presentations continued in three working groups.
The first group discussed philosophical questions regarding the design and impact of data–model comparison studies, summarized in Figure 1. Among these were what insights into climate can be gained from data–model comparison, and how both sources of information can be leveraged. The importance of formulating a clear hypothesis prior to the comparison was stressed, as well as the need to explain experimental choices and assumptions such that the scope and limitations of the respective analysis are clearly defined. Consequently, techniques for a rigorous treatment of uncertainties are required and due to various uncertainties on both sides, data and models are not necessarily expected to "agree".
 |
|
Figure 1: Key components and challenges of data–model comparison. The relevant tools, variables, intercomparison projects, and challenges (in orange) are illustrated with respect to the targeted time ranges. The workshop specifically addressed the overarching question of how paleoclimatology can contribute to solving research questions on future climate scenarios. |
The second group formulated a research project based on the recommendations from the first group. Inspired by the keynote talks, the group decided to study the temperature–hydroclimate relationship in the tropics and test the hypothesis of positive covariability between the two. The participants identified suitable databases (e.g. PAGES2k Consortium 2017; Konecky et al. 2019; Comas-Bru et al. 2020) for a multi-archive and multi-proxy approach. Comparison against isotope-enabled simulations (e.g. Bühler et al. 2021) is planned. Key questions that the group identified include whether emerging isotope-enabled simulations facilitate more robust data–model comparisons, and how multiple archives and simulations can be used to understand the underlying mechanisms controlling the covariability of hydroclimate and temperature.
The third group started with the conundrum of reported agreement of global mean temperature variability in models and proxies on decadal-to-centennial scales (e.g. Neukom et al. 2019), whereas reconstructed local surface temperature variability is higher than in simulations (e.g. Laepple and Huybers 2014). The group reviewed the literature, with a focus on the spatial and temporal scales of interest. Finally, the group collected and assessed potential reasons to explain the conundrum, including effects from an overestimation of spatially uncorrelated variability in temperature reconstructions, misspecification of the spatial correlation structures in models, and the suppression of variability by climate field reconstruction methods. The group plans to expand the literature review and develop research protocols to quantify the contributions of identified potential explanations.
For most participants, the workshop was the first experience with a hybrid conference format, and the feedback was quite positive. We emphasize the importance of an appropriate technical infrastructure on site and the prior set-up of a clear workshop structure. The use of a virtual communication platform and shared working documents helped to connect virtual and on-site participants.
Affiliations
1Institute of Environmental Physics, Heidelberg University, Germany
2Department of Geography, University College London, UK
3Institute for Geosciences, University of Bonn, Germany
4Potsdam Institute for Climate Impact Research, Germany
5Geo- und Umweltforschungszentrum, Tübingen University
contact
Nils Weitzel: nweitzeliup.uni-heidelberg.de
references
Bühler J et al. (2021) Clim Past 17: 985-1004
Comas-Bru L et al. (2020) Earth Syst Sci Data 12: 2579-2606
Konecky BL et al. (2019) Earth Syst Sci Data 12: 2261-2288
Laepple T, Huybers P (2014) Proc Natl Acad Sci USA 111: 16682-16687
Marie-France Loutre![]() 1, P.A. Gell
1, P.A. Gell![]() 2, K.J. Meissner
2, K.J. Meissner![]() 3 and B. Vannière
3 and B. Vannière![]() 4
4
Online, 15 June 2021
PAGES organized a session during the hybrid Sustainability Research & Innovation Congress 2021 (SRI2021; pastglobalchanges.org/calendar/26956), a joint initiative of Future Earth and the Belmont Forum, which was led by the Commonwealth Scientific and Industrial Research Organisation (CSIRO) and Future Earth Australia.
International reports on climate, biodiversity and ecosystems, such as the Intergovernmental Panel on Climate Change (IPCC) and Intergovernmental Platform on Biodiversity and Ecosystem Services (IPBES), describe the unprecedented changes that the Earth system has experienced over the last few decades. While the data are clear, it is difficult for a human being to identify these changes and their potential consequences—partly because our individual memory is short, and partly because on a daily or annual basis we experience much larger changes and quickly adapt and get used to new conditions. In other words, we struggle to identify the baselines of changes and how these baselines shift with time. Therefore, it is important to identify reference points for the most recent changes, or at least to be aware of how humans are shaping the climate and ecosystems.
Human-induced climate change is altering some of the processes that underlie modes of climate variability that can have major impacts on societies. Short observational records make it difficult to understand the full range of natural climate variability and to robustly detect recent changes in these modes of variability. Nerilie Abram reviewed paleoclimate evidence for changes in the El Niño-Southern Oscillation, Indian Ocean Dipole and Southern Annular Mode during the last millennium. She discussed how unusual some of these phenomena have been over recent years and gave perspectives on the likely future of these systems (Fig. 1a).
 |
|
Figure 1: Illustration of the four topics discussed during this session. (A) Modes of climate variability; (B) human impacts on aquatic ecosystems; (C) flood and (D) fire hazards. Image credits: (A) Australian Government, Bureau of Meteorology; (B) Peter Gell; (C) Meteoschweiz (Gasthaus zur Krone, Basel; Staatsarchiv Basel-Stadt, BILD13, 323); (D) Matthew Abbot (Lake Conjola, 31 Dec 2019; matthewabbott.com.au/PHOTOJOURNALISM/Black-Summer/1). |
Michael Reid described the deep history of human impacts on aquatic ecosystems. He highlighted how paleoecological reconstructions can provide avenues to determine impacts on ecosystems caused by human activities, including activities that took place long before the establishment of systematic ecological monitoring. He stressed the importance of understanding the relative and interactive effects of multiple stressors and showed, with an interesting example from the Murray-Darling Basin in Australia, the need to address all stressors, not just the most recent ones (Fig. 1b).
Threats to property and people linked to floods are becoming more and more important, in particular because of increasing population densities in areas prone to flooding. Furthermore, the magnitude, frequency, and timing are changing, thus exacerbating the issue. Extreme flooding can be documented through historical, botanical, and geological records. Bruno Wilhelm reviewed how these archives record floods and the information they can provide (Fig. 1c). He then showed how the flood risk assessment can be improved with these records and discussed avenues for improvement. Flood management and mitigation plans should take this information into account.
Simon G. Haberle provided a long-term perspective on the 2019-2020 Australian Bushfire Crisis. The paleoecological records from pollen and charcoal show an increase in fire activity during times of past climate change – particularly during transitions from cooler to warmer climates. The presence of humans on the Australian landscape for at least the last 65,000 years appears to have mediated past climate impacts on fire regimes and fire sensitive ecosystems through Aboriginal fire management practices. The cessation of these landscape management practices across much of Australia with the start of European colonization more than 200 years ago induce a shift from low but persistent levels of burning to a much more variable fire regime pattern (Fig. 1d).
The session concluded with a fruitful panel discussion. The importance of past climate and ecosystem reconstructions was discussed, especially in light of their capacity to improve risk assessments related to current and future changes. The need to improve communication channels between past global change experts and policy makers or other end users, such as insurance companies, was also highlighted. Overall, this session successfully brought together experts from different areas of the climate system and ecosystems to discuss changes, transitions, and resilience of vital systems upon which our well-being depends.
Affiliations
1Past Global Changes International Project Office, Bern, Switzerland
2Faculty of Science & Technology, Federation University Australia, Ballarat, Australia
3Climate Change Research Centre, University of New South Wales, Sydney, Australia
4Chrono-environnement, CNRS - Université de Franche-Comté, Besançon, France
contact
Marie-France Loutre: marie-france.loutrepages.unibe.ch